
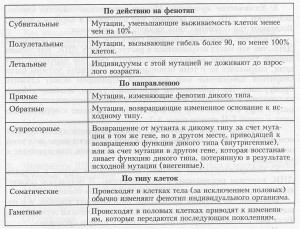
Хромосомные мутации (перестройки, или аберрации) — это изменения в структуре хромосом, которые можно выявить и изучить под световым микроскопом.








Известны перестройки разных типов (рис. 3.13):
- нехватка, или дефишенси, — потеря концевых участков хромосомы;
- делеция — выпадение участка хромосомы в средней ее части;
- дупликация — двух- или многократное повторение генов, локализованных в определенном участке хромосомы;
- инверсия — поворот участка хромосомы на 180°, в результате чего в этом участке гены расположены в последовательности, обратной по сравнению с обычной;
- транслокация — изменение положения какого-либо участка хромосомы в хромосомном наборе. К наиболее распространенному типу транслокаций относятся реципрокные, при которых происходит обмен участками между двумя негомологичными хромосомами. Участок хромосомы может изменить свое положение и без реципрокного обмена, оставаясь в той же хромосоме или включаясь в какую-то другую.
При дефишенси, делециях и дупликациях изменяется количество генетического материала. Степень фенотипического изменения зависит от того, насколько велики соответствующие участки хромосом и содержат ли они важные гены. Примеры дефишенси известны у многих организмов, включая человека. Тяжелое наследственное заболевание —синдром «кошачьего крика» (назван так по характеру звуков, издаваемых больными младенцами), обусловлен гетерозиготностью по дефишенси в 5-й хромосоме. Этот синдром сопровождается сильным нарушением роста и умственной отсталостью. Обычно дети с таким синдромом рано умирают, но некоторые доживают до зрелого возраста.
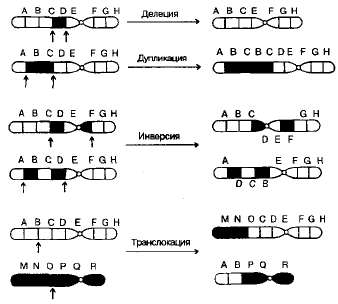
3.13. Хромосомные перестройки, изменяющие расположение генов в хромосомах.
Геномные мутации — изменение числа хромосом в геноме клеток организма. Это явление происходит в двух направлениях: в сторону увеличения числа целых гаплоидных наборов (полиплоидия) и в сторону потери или включения отдельных хромосом (анеуплоидия).
Полиплоидия — кратное увеличение гаплоидного набора хромосом. Клетки с разным числом гаплоидных наборов хромосом называются триплоидными (Зn), тетраплоидными (4n), гексанло-идными (6n), октаплоидными (8n) и т. д.
Чаще всего полиплоиды образуются при нарушении порядка расхождения хромосом к полюсам клетки при мейозе или митозе. Это может быть вызвано действием физических и химических факторов. Химические вещества типа колхицина подавляют образование митотического веретена в клетках, приступивших к делению, в результате чего удвоенные хромосомы не расходятся и клетка оказывается тетрагшоидной.
Для многих растений известны так называемые полиплоидные ряды. Они включают формы от 2 до 10n и более. Например, полиплоидный ряд из наборов в 12, 24, 36, 48, 60, 72, 96, 108 и 144 хромосомы составляют представители рода паслен (Solanum). Род пшеница (Triticum) представляет ряд, члены которого имеют 34, 28 и 42 хромосомы.
Полиплоидия приводит к изменению признаков организма и поэтому является важным источником изменчивости в эволюции и селекции, особенно у растений. Это связано с тем, что у растительных организмов весьма широко распространены гермафродитизм (самоопыление), апомиксис (партеногенез) и вегетативное размножение. Поэтому около трети видов растений, распространенных на нашей планете, — полиплоиды, а в резко континентальных условиях высокогорного Памира произрастает до 85% полиплоидов. Почти все культурные растения тоже полиплоиды, у которых, в отличие от их диких сородичей, более крупные цветки, плоды и семена, а в запасающих органах (стебель, клубни) накапливается больше питательных веществ. Полиплоиды легче приспосабливаются к неблагоприятным условиям жизни, легче переносят низкие температуры и засуху. Именно поэтому они широко распространены в северных и высокогорных районах.
В основе резкого увеличения продуктивности полиплоидных форм культурных растений лежит явление полимерии (см. § 3.3).
Анеуплоидия, или гетероплодия, — явление, при котором клетки организма содержат измененное число хромосом, не кратное гаплоидному набору. Анеуплоиды возникают тогда, когда не расходятся или теряются отдельные гомологичные хромосомы в митозе и мейозе. В результате нерасхождения хромосом при гамето-генезе могут возникать половые клетки с лишними хромосомами, и тогда при последующем слиянии с нормальными гаплоидными гаметами они образуют зиготу 2n + 1 (трисомик) по определенной хромосоме. Если в гамете оказалось меньше на одну хромосому, то последующее оплодотворение приводит к образованию зиготы 1n - 1 (моносомик) по какой-либо из хромосом. Кроме того, встречаются формы 2n - 2, или нуллисомики, так как отсутствует пара гомологичных хромосом, и 2n + х, или полисомики.
Анеуплоиды встречаются как у растений и животных, так и у человека. Анеуплоидные растения обладают низкой жизнеспособностью и плодовитостью, а у человека это явление нередко приводит к бесплодию и в этих случаях не наследуется. У детей, родившихся от матерей старше 38 лет, вероятность анеуплоидии повышена (до 2,5%). Кроме того, случаи анеуплоидии у человека вызывают хромосомные болезни.
У раздельнополых животных как в естественных, так и в искусственных условиях полиплоидия встречается крайне редко. Это обусловлено тем, что полиплоидия, вызывая изменение соотношения половых хромосом и аутосом, приводит к нарушению конъюгации гомологичных хромосом и тем самым затрудняет определение пола. В результате такие формы оказываются бесплодными и маложизнеспособными.
Спонтанные и индуцированные мутации. Спонтанными называют мутации, возникающие под влиянием неизвестных природных факторов, чаще всего как результат ошибок при воспроизведении генетического материала (ДНК или РНК). Частота спонтанного мутирования у каждого вида генетически обусловлена и поддерживается на определенном уровне.
Индуцированный мутагенез — это искусственное получение мутаций с помощью физических и химических мутагенов. Резкое увеличение частоты мутаций (в сотни раз) происходит под воздействием всех видов ионизирующих излучений (гамма- и рентгеновские лучи, протоны, нейтроны и др.), ультрафиолетового излучения, высоких и низких температур. К химическим мутагенам относятся такие вещества, как формалин, азотистый иприт, колхицин, кофеин, некоторые компоненты табака, лекарственных препаратов, пищевых консервантов и пестицидов. Биологическими мутагенами являются вирусы и токсины ряда плесневых грибов.
В настоящее время ведутся работы по созданию методов направленного воздействия различных мутагенов на конкретные гены. Такие исследования очень важны, поскольку искусственное получение мутаций нужных генов может иметь большое практическое значение для селекции растений, животных и микроорганизмов.
Закон гомологических рядов в наследственной изменчивости. Крупнейшим обобщением работ по изучению изменчивости в начале XX в. стал закон гомологических рядов в наследственной изменчивости. Он был сформулирован выдающимся русским ученым Н. И. Вавиловым в 1920 г. Сущность закона заключается в следующем: виды и роды, генетически близкие, связанные друг с другом единством происхождения, характеризуются сходными рядами наследственной изменчивости. Зная, какие формы изменчивости встречаются у одного вида, можно предвидеть нахождение аналогичных форм у родственного ему вида.
В основе закона гомологических рядов фенотипической изменчивости у родственных видов лежит представление о единстве их происхождения от одного предка в процессе естественного отбора. Поскольку общие предки имели специфический набор генов, то их потомки должны обладать примерно таким же набором.
Более того, у родственных видов, имеющих общее происхождение, возникают и сходные мутации. Это означает, что у представителей разных семейств и классов растений и животных со сходным набором генов можно встретить параллелизм — гомологические ряды мутаций по морфологическим, физиологическим и биохимическим признакам и свойствам. Так, у разных классов позвоночных встречаются сходные мутации: альбинизм и отсутствие перьев у птиц, альбинизм и бесшерстность у млекопитающих, гемофилия у многих млекопитающих и человека. У растений наследственная изменчивость отмечена по таким признакам, как пленчатое или голое зерно, остистый или безостый колос и др.
Закон гомологических рядов, отражая общую закономерность мутационного процесса и формообразования организмов, представляет широкие возможности для его практического использования в сельскохозяйственном производстве, селекции, медицине. Знание характера изменчивости нескольких родственныхх видов дает возможность поиска признака, который отсутствует у одного из них, но характерен для других. Таким путем были собраны и изучены голозерные формы злаков, односемянные сорта сахарной свеклы, не нуждающиеся в прорывке, что особенно важно при механизированной обработке почв. Медицинская наука в качестве моделей для изучения болезней человека получила возможность использовать животных с гомологическими заболеваниями: это сахарный диабет крыс; врожденная глухота мышей, собак, морских свинок; катаракта глаз мышей, крыс, собак и др.
Закон гомологических рядов позволяет также предвидеть возможность появления мутаций, еще неизвестных науке, которые могут использоваться в селекции для создания новых ценных для хозяйства форм.
Типы мутаций
Вполне вероятно, что у плодовых мушек, которых облучал Мюллер, появлялось гораздо больше мутаций, чем он смог обнаружить. По определению, мутация — это любое изменение в ДНК. Это значит, что мутации могут происходить в геноме где угодно. А поскольку большую часть генома занимает «мусорная» ДНК, ничего не кодирующая, большинство мутаций остаются незамеченными.
Мутации изменяют физические свойства организма (признаки), только если они изменяют последовательность ДНК внутри гена (рис. 7.1).
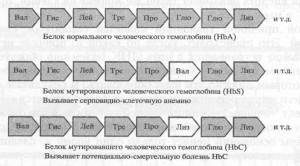
Рис. 7.1. Эти три аминокислотные последовательности показывают, как маленькие изменения могут приводить к большому эффекту. Начало одной из аминокислотных цепей в нормальном белке приведено в верхнем ряду. Ниже аминокислотная цепь ненормального варианта белка гемоглобина: валин замешен на глютаминовую кислоту в шестом положении. Эта единственная замена, приводящая к мутации кодона ГАА в кодон ГУА, является причиной серповидно-клеточной анемии, выражающейся в ряде симптомов: от слабой анемии (если у индивидуума остается нормальная копия мутировавшего гена) до смерти (если у индивидуума две мутировавшие копии гена)
Хотя Мюллер индуцировал мутации у плодовых мушек, подвергая их высоким дозам облучения, мутации случаются в организме все время. Иногда это просто ошибки нормальных процессов, происходящих в клетке, а иногда — результат воздействия окружающей среды. Такие спонтанные мутации встречаются с частотами, характерными для определенного организма, иногда называемыми спонтанным фоном.
Наиболее часто случаются точковые мутации, которые изменяют всего одну пару оснований в нормальной последовательности ДНК. Их можно получить двумя путями:
1. ДНК химически модифицируется, так что одно из оснований меняется на другое.
2. Репликация ДНК работает с ошибками, вставляя ошибочное основание в цепь при синтезе ДНК.
Какова бы ни была причина их появления, точковые мутации можно разделить на два типа:
1. Транзиции. Наиболее часто встречающийся тип мутаций. При транзиции один пиримидин замещается другим пиримидином или один пурин замещается другим пурином: например, пара Г-Ц становится парой А-Т, или наоборот.
2. Трансверзии. Более редкий тип мутаций. Пурин замещается пиримидином или наоборот: например, пара А-Т становится парой Т-А или Ц-Г.
Азотистая кислота — это мутаген, который вызывает транзиции. Она конвертирует цитозин в урацил. Цитозин обычно дает пару с гуанином, но урацил — с аденином. В результате пара Ц-Г становится парой Т-А, когда А спаривается с Т в следующей репликации. Азотистая кислота оказывает такой же эффект на аденин, превращая пару А-Т в пару Ц-Г.
Другой причиной транзиций является ошибочное спаривание оснований. Это происходит, когда по какой-то причине неправильное основание встраивается в нить ДНК, затем оно образует пару с неправильным партнером (некомплементарным основанием) вместо того, с которым должно эту пару образовать. В результате во время следующего цикла репликации пара полностью меняется.
Эффект точковых мутаций зависит от того, в каком месте последовательности оснований они образуются. Поскольку изменение одной пары оснований меняет только один кодон и, следовательно, одну аминокислоту, получающийся в результате белок может быть поврежден, но может, несмотря на повреждение, сохранить часть нормальной активности.
Гораздо сильнее, чем точковые мутации, повреждают ДНК мутации сдвига рамки. Вспомните, что генетическая последовательность оснований (секвенс) считывается как последовательность неперекрывающихся триплетов (трех оснований). Это значит, что существует три пути прочтения (рамки считывания) последовательности оснований, зависящих от точки начала прочтения. Если мутация убирает или встраивает лишнее основание, она вызывает сдвиг рамки, и вся последовательность оснований прочитывается неправильно. Это значит, что изменится вся последовательность аминокислот, а получающийся белок, с большой долей вероятности, будет полностью неработающим.
Мутации сдвига рамки вызываются акридинами, химическими веществами, которые связываются с ДНК и настолько изменяют ее структуру, что основания могут быть добавлены или убраны из ДНК во время ее репликации. Эффект таких мутаций зависит от места последовательности оснований, в котором произойдет вставка (инсерция) или выпадение (делеция) оснований, а также от их взаимного расположения в образующейся последовательности (рис. 7.2).
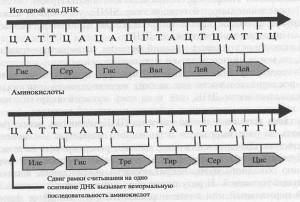
Рис. 7.2. Один из способов, которым мутация сдвига рамки может влиять на считывание последовательности оснований ДНК
Еще одним типом мутаций является встраивание (инсерция) длинных фрагментов дополнительного генетического материала в геном. Встраиваются транспозирующиеся (мобильные генетические) элементы, или транспозоны, — последовательности, которые могут перемещаться из одного места ДНК в другое. Впервые транспозоны были открыты генетиком Барбарой МакКлинток (Barbara McClintock) в 1950-е годы. Это короткие элементы ДНК, которые из одной точки генома могут перепрыгнуть в другую (поэтому их часто называют «прыгающими генами»). Иногда они прихватывают с собой расположенные рядом последовательности ДНК. Обычно транспозоны состоят из одного или нескольких генов, один из которых представляет собой ген фермента транспозазы. Этот фермент требуется транспозонам для перемещения из одного места ДНК в другое внутри клетки.
Существуют также ретротранспозоны, или ретропозоны, которые сами передвигаться не могут. Вместо этого они используют свою мРНК. Она сначала копируется в ДНК, а последняя вставляется в другую точку генома. Ретротранспозоны родственны ретровирусам.
Если транспозон встраивается в ген, кодирующая последовательность оснований нарушается, и ген в большинстве случаев выключается. Транспозоны также могут нести сигналы для окончания транскрипции или трансляции, которые эффективно блокируют выражение других генов, расположенных вслед за ними. Такой эффект называется полярной мутацией.
Ретротранспозоны типичны для геномов млекопитающих. Фактически, около 40% генома состоит из таких последовательностей. Это одна из причин, по которой геном содержит так много «мусорной» ДНК. Ретротранспозоны могут быть SINE (короткими промежуточными элементами) длиной в несколько сот пар оснований пли LINE (длинными промежуточными элементами) длиной от 3000 до 8000 пар оснований. Например, человеческий геном содержит около 300 тыс. последовательностей одного типа SINE, у которых, кажется, нет другой функции, кроме саморепликации. Данные элементы также называются «эгоистической» ДНК.
В отличие от точковых мутаций мутации, вызываемые транспозонами, не могут индуцироваться мутагенами.
Точковые мутации могут ревертировать, возвращаться к исходной последовательности как за счет восстановления оригинальной последовательности ДНК, так и за счет мутаций в других местах гена, которые компенсируют действие первичной мутации.
Вставка дополнительного элемента ДНК, очевидно, может ревертировать за счет вырезания вставленного материала — точечного исключения. Делеция части гена, однако, ревертировать не может.
Мутации могут происходить в других генах, приводя к формированию обходного пути, исправляющего повреждение, вызванное начальной мутацией. В результате образуется двойной мутант, имеющий нормальный или почти нормальный фенотип. Этот феномен называется супрессией, бывающей двух типов: внегенной и внутригенной.
Внегенная супрессорная мутация подавляет действие мутации, расположенной в другом гене, иногда за счет изменения физиологических условий, при которых белок, кодируемый супрессируемым мутантом, может функционировать вновь. Бывает, что такая мутация меняет аминокислотную последовательность мутантного белка.
Внутригенная супрессорная мутация подавляет эффект мутации в гене, где она расположена, иногда восстанавливая рамку считывания, нарушенную мутацией сдвига рамки. В некоторых случаях мутация изменяет аминокислоты в сайте, который компенсирует изменение аминокислоты, вызванное первичной мутацией. Феномен также называется реверсией во втором сайте.
Не все последовательности оснований в гене подвержены мутациям в одинаковой мере. Мутации имеют тенденцию группироваться вокруг горячих точек в последовательности гена — местах, где вероятность образования мутаций в 10 или 100 раз выше, чем ожидаемая при случайном распределении. Расположение этих горячих точек различно для разных типов мутаций и мутагенов, индуцирующих их.
В бактерии E . coli, например, горячие точки встречаются там, где расположены модифицированные основания, называемые 5-метил-цитозином. Это основание иногда подвергается таутомерному сдвигу — перестройке атома водорода. В результате Г спаривается с Т вместо Ц, а после репликации образуется пара дикого типа Г-Ц и мутантная пара А-Т (в генетике диким типом называются последовательности ДНК, которые обычно встречаются в природе).
Многие мутации не дают видимого эффекта. Они называются молчащими мутациями. Иногда мутация молчит, потому что изменения не влияют на продукцию аминокислот, а иногда — поскольку, несмотря на замену аминокислоты в белке, новая аминокислота не влияет на его функцию. Это называется нейтральной заменой.
Мутация, выключающая или изменяющая функцию гена, называется прямой мутацией. Мутация, которая реактивирует или восстанавливает функцию гена за счет реверсии начальной мутации или за счет открытия обходного пути (как при реверсии во втором сайте, описанной выше), называется обратной мутацией.
Как видите, есть много различных способов классифицировать мутации, и одна и та же мутация может относиться к различным типам. Данные табл. 7.1 могут внести ясность в характеристику мутаций.
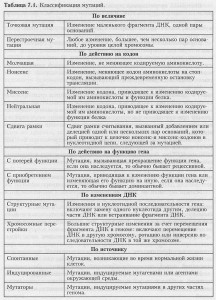
Классификация мутаций
Классификация мутаций (продолжение)
30. Генетика и эволюционная теория.
Разработанная Дарвином эволюционная теория основывается на следующих факторах: изменчивости, наследственности, борьбе за существование и естественном отборе. Главное значение как материал для отбора имеет, по Дарвину, наследственная изменчивость. В настоящее время эти положения глубоко разрабатываются.
В свете современных научных данных можно утверждать, что основу дарвиновской наследственной изменчивости составляют мутации. Их и следует рассматривать как основной первичный материал для эволюционного процесса. Особи, несущие одни мутации, скрещиваются с особями, имеющими другие мутации. Получаются новые сочетания генов, новые генотипы. Эта изменчивость и дает первичный материал для естественного отбора, ведущего к образованию новых видов.
Генетика популяций. Основной формой существования вида являются популяции. Для того чтобы конкретно представить начальные этапы эволюции, важно изучить те генетические процессы, которые протекают в популяциях. Действие генетических законов в популяциях представляет предмет исследования особого раздела генетики, называемого генетикой популяций и имеющего большое значение для эволюционной теории. Познакомимся с некоторыми положениями этой области генетики.
Допустим, что в популяции происходит свободное скрещивание доминантной и рецессивной форм, отличающихся по одной паре аллелей: AA и aa. В первом поколении все гибриды будут гетерозиготны, а далее в F2 и последующих поколениях пойдет расщепление. В F2 будет следующее соотношение генотипов: 1AA+2Aa+1aa. Каковы же будут в последующих поколениях соотношения гомо- и гетерозигот при свободном скрещивании? Это легко рассчитать, если определить, сколько гамет и каких получим от различных генотипов:
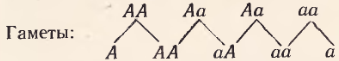
Простой подсчет показывает, что гамет, несущих ген A и несущих ген a (аллельные гены), будет поровну. Следовательно, при образовании зигот возникнут следующие комбинации:

То же соотношение повторится и в поколениях.
Из поколения в поколение при свободном скрещивании относительные частоты генов (и соответственно гомо- и гетерозигот) не меняются. Эта закономерность по имени установивших ее в 1908 г. ученых носит название закона Харди – Вайнберга. Закон справедлив, однако лишь при соблюдении следующих условий: популяция должна быть достаточно велика, чтобы обеспечить возможность случайного сочетания генов; должен отсутствовать отбор, благоприятствующий или неблагоприятствующий определенным генам; не должно возникать новых мутаций; не должна происходить миграция особей с иными генотипами из соседних популяций данного вида. В существующих в природе популяциях эти условия не соблюдаются, так как действует естественный отбор, происходят мутации и миграции особей. Это приводит к нарушению равновесия генов в популяциях.
Постоянно протекающий мутационный процесс и свободное скрещивание приводят к тому, что в пределах популяции накапливается большое количество внешне непроявляющихся наследственных изменений (подавляющее большинство возникающих мутаций рецессивно) Эти важные для понимания хода начальных стадий эволюции факты были установлены советским ученым С. С. Четвериковым. Генетические исследования природных популяций растений и животных показали, что при относительной фенотипической однородности они насыщены разнообразными рецессивными мутациями. Хромосомы, в которых возникли мутации, в результате удвоения при делении клетки постепенно распространяются среди популяций. Мутации не проявляются фенотипически до тех пор, пока остаются гетерозиготными. По достижении достаточно высокой концентрации мутаций становится вероятным скрещивание особей, несущих аллельные рецессивные гены. В этих случаях мутации проявятся фенотипически и подпадут под прямой контроль естественного отбора. Таким образом, каждый вид и каждая его популяция представляют сложную гетерозиготную систему, таящую в себе, по выражению академика И. И. Шмальгаузена, «резерв наследственной изменчивости», который может быть «мобилизован» через естественный отбор при изменении условий существования популяций. Для каждой популяции характерен свой генофонд. Генофонд это совокупность генов популяции, вида или иной систематической группы.
Каждая популяция как бы таит в своих недрах возможности для быстрого изменения в соответствии с направлением естественного отбора.
Формы естественного отбора. Знакомство с генетикой позволяет нам углубить и конкретизировать вопрос о разных формах естественного отбора, протекающего в природе. В разных условиях среды действие естественного отбора носит различный характер. Предположим, что создались условия, при которых отдельные возникающие наследственные уклонения полезны. В этом случае действие отбора будет направлено в одну определенную сторону. Это приведет к постепенному изменению фенотипа, к смене нормы реакции в одном определенном направлении 119. Такая форма отбора носит название движущего отбора Приведем пример. Близ индустриальных центров в воздухе много копоти, дыма. Стволы берез приобретают грязно-коричневый оттенок. У живущей на березе бабочки березовой пяденицы иногда появляются темноокрашенные мутанты. В обычных условиях сельской местности они отметаются отбором, так как темная окраска делает бабочек заметными на фоне белой коры березы, и их поедают птицы. Иное дело – на загрязненной дымом березе. В этих условиях темные пяденицы становятся менее заметными и естественный отбор их сохраняет. Фактором, осуществляющим этот отбор, преимущественно служат птицы, поедающие бабочек. При большой интенсивности отбора через короткий промежуток времени возникает популяция, характеризующаяся темной окраской. Например, в окрестностях города Манчестера темная форма березовой пяденицы вытеснила светлую форму примерно за 20 лет. Движущий отбор играет основную роль в эволюции, в развитии приспособлений. Так, например, протекала эволюция лошади – от пятипалой конечности к однопалой; и так же шло образование бескрылых островных форм насекомых, и т. п.
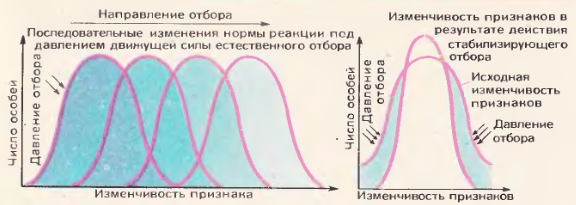 Рис. 119. Форма естественного отбора
Рис. 119. Форма естественного отбора
Наряду с движущим естественным отбором в природе широко осуществляется и другая его форма – стабилизирующий отбор. У видов, живущих в относительно постоянных условиях, возникающие изменения могут быть неблагоприятными. В таких случаях сохраняются мутации, ведущие к меньшей изменчивости данного признака, и отсекаются мутации, определяющие более широкую изменчивость Вот пример действия стабилизирующего отбора. У опыляемых насекомыми растений малой изменчивостью характеризуются части цветка. Это связано с тем, что пропорции цветка приспособлены к размерам опыляющих их насекомых и широкая изменчивость здесь отразилась бы неблагоприятно на ходе опыления. Под действием стабилизирующего отбора устойчиво закрепились пропорции и размеры частей цветка.
Другой пример действия стабилизирующего отбора приведен выше. После снегопада и бури, прошедшей в Англии, выжили преимущественно воробьи со средней длиною крыла. Длиннокрылые и короткокрылые погибали в большем количестве. Отбор в данном случае как бы «сохраняет норму». Если условия жизни вида долгое время не меняются, то стабилизирующий отбор способствует не изменению, а сохранению признаков вида, приспособленных к данным условиям. Если же условия существования вида изменятся, то ведущую роль приобретают движущие формы отбора, приводящие к изменению вида, к видообразованию.
Движущая и стабилизирующая формы отбора в природе тесно связаны друг с другом. Движущий отбор преобразует виды в меняющихся условиях окружающей среды. Стабилизирующий отбор закрепляет полезные формы в относительно постоянных условиях среды [120].
 Рис. 120. Пример действия стабилизирующего отбора.
Рис. 120. Пример действия стабилизирующего отбора.
В относительно постоянных условиях среды стабилизующий отбор способствует сохранению полезных виду признаков (плавники, обтекаемая форма, пропорция тела – все приспособлено к условиям существования в водной среде).
Генетический анализ популяций позволяет значительно углубить и уточнить наши знания о характере изменчивости организмов в природе н конкретно уяснить механизм действия естественного отбора как основного фактора микроэволюции и видообразования.
31. селекция животных, методы и достижения.
Селекция животных
Новые породы животных получают на основе наследственной изменчивости путем отбора. Как для растений, так и для животных необходимы определенные условия среды, которые способствуют наиболее полному проявлению генетических задатков и развитию желаемых качеств. Сущность селекции животных состоит в сохранении, усилении и комбинировании у потомства ценных и устранении нежелательных качеств. Работа по созданию, поддержанию и усовершенствованию пород включает ряд методов разведения и организационных мероприятий, которые в совокупности представляют племенное дело. Оно имеет свои особенности. Домашние животные в отличие от растений размножаются только половым путем, а половая зрелость у некоторых из них наступает через несколько лет. Самка рождает одного или нескольких детенышей, что замедляет процесс селекции.
Одомашнивание животных — первый этап в селекции. Оно началось 10—12 тыс. лет до н. э., а основная часть домашних животных появилась в неолите, 5—6 тыс. лет назад. Эта работа продолжается и сейчас (например, одомашнивание тувинского яка, антилопы канны, дающей ценное по питательности и лечебным свойствам молоко). Одомашнивание резко повышает изменчивость организмов и создает благоприятные условия для искусственного отбора. В настоящее время усиленно развивается новая отрасль сельского хозяйства — пушное звероводство. Влияние приручения на изменчивость пушных зверей (лисы, песца, норки) многие годы изучал академик Д. К. Беляев.
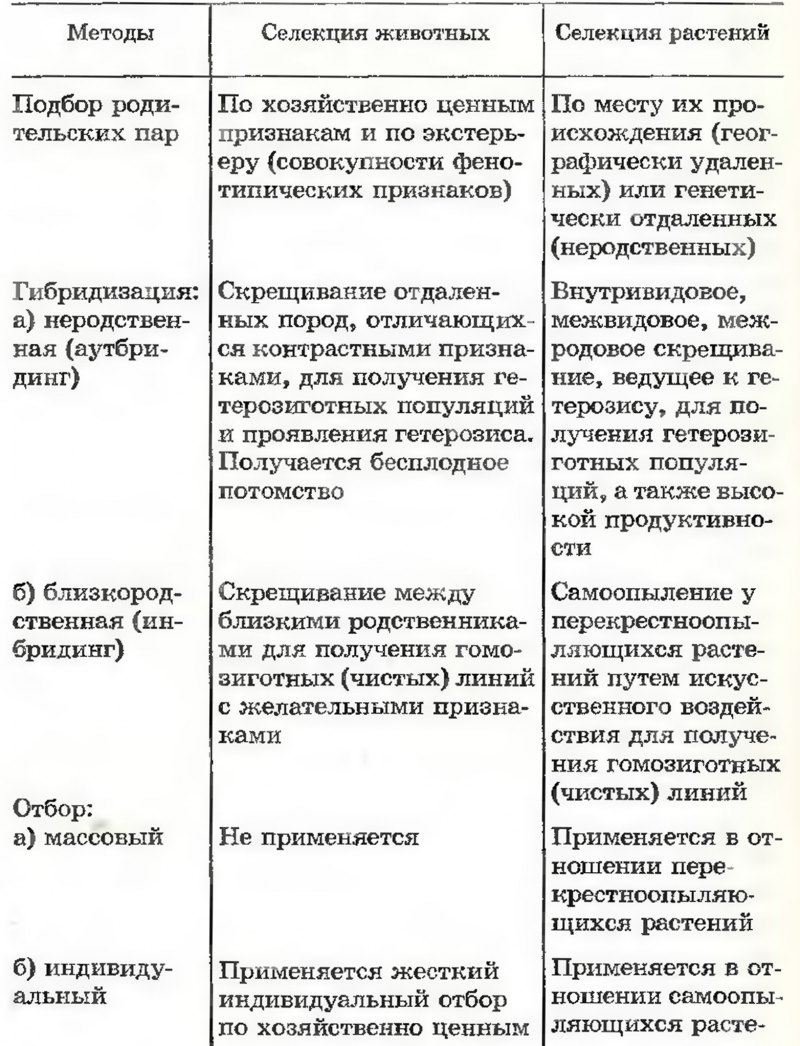
Методы селекции в животноводстве. Отбор родительских пар совершается в зависимости от цели, которую поставил селекционер (повышение удоев, жирности молока, качества мяса и т. д.). Разводимых животных оценивают по фенотипу, происхождению и по качеству их потомства. Поэтому необходимо хорошо знать их родословную.
Все сельскохозяйственные животные раздельнополы. В то же время многие виды ценной животноводческой продукции создаются животными только одного пола (молоко, яйца). Поэтому оценка животных другого пола может быть осуществлена по их родословной и по качеству их потомства. Так, племенные качества быка-производителя могут быть оценены по молочной продуктивности его предков по материнской линии, его сестер и особенно его дочерей.
Основной способ наследственного разнообразия при селекционной работе с животными — скрещивание. Оно может быть родственным и неродственным. Родственное скрещивание — инбридинг — между братьями и сестрами или между родителями и потомством применяется, когда селекционер хочет большинство генов данной породы привести в гомозиготное состояние. Такое скрещивание похоже на самоопыление и ведет к гомозиготности. Оно сопровождается строгим отбором по необходимым хозяйственным качествам и чаще всего приводит к ослаблению животных, уменьшению устойчивости к действию внешних факторов, к заболеваниям и т. д. Для устранения этих неблагоприятных последствий используется скрещивание различных линий и пород. Ценность родственного скрещивания заключается в том, что позволяет закрепить в породе полезные хозяйственные качества. Неродственное скрещивание в границах породы или между породами, сопровождаемое строгим отбором, ведет к поддержанию полезных качеств и к усилению их в следующих поколениях.
Как у растений, так и у животных наблюдается явление гетерозиса. Его сущность состоит в том, что в первом поколении гибриды имеют повышенную жизнеспособность и усиленное развитие. При последующих скрещиваниях гибридов между собой эти качества ослабевают (по-видимому, вследствие вы-щепления гомозигот). Гетерозис применяют в овцеводстве, молочном скотоводстве, свиноводстве. Примером особенно эффективного использования гетерозиса служит выведение гетерозисных цыплят — бройлерное производство. Оно широко применяется в птицеводстве многих стран.
Для разработки научных методов селекции сельскохозяйственных животных большое значение имела селекционная работа академика М. Ф. Иванова. В условиях юга Украины им была создана высокопродуктивная порода свиней Белая украинская. Начало ей положили две породы: местная, хорошо приспособленная к климатическим условиям, но с низкой продуктивностью, и Английская белая — высокопродуктивная, но совершенно непригодная к содержанию в условиях юга Украины. Скрестив эти породы между собой, М. Ф. Иванов в первых гибридных поколениях использовал инбридинг. Работа сопровождалась жестким отбором животных, выращиваемых в условиях, для которых создавалась порода. В результате в новой породе высокая продуктивность была соединена с хорошей приспособленностью к местным условиям. Порода овец Асканийская рамбулье — другой пример высокопродуктивной породы, созданной М. Ф. Ивановым.
Отдаленная гибридизация домашних животных. Работая в Институте акклиматизации и гибридизации животных в Аскания-Нова, М. Ф. Иванов много сделал для применения отдаленной гибридизации в селекционных целях. В настоящее время в нашей стране такие работы проводятся в широком масштабе. В хозяйственном отношении особенно ценны скрещивания между крупным рогатым скотом и яками, крупным рогатым скотом и зебу, домашними породами овец и диким бараном архаром. В результате были получены животные, сочетающие ценные качества исходных видов. Однако часто отдаленная гибридизация приводит к бесплодию гибридов вследствие нарушения нормального течения гаметогенеза. Примером может служить мул — гибрид лошади и осла. И хотя мулы совершенно не дают потомства, во многих странах их широко используют из-за высокой выносливости и долговечности.
Примерами достижений селекции животных в нашей стране могут служить высокопородные, приспособленные к природе Сибири овцы, пригодная для выращивания в условиях промышленного комплекса порода свиней Сибирский ландрас, хорошо приспособленные к стойловому содержанию популяции крупного рогатого скота мясо-молочной направленности и др.
Основные методы селекции
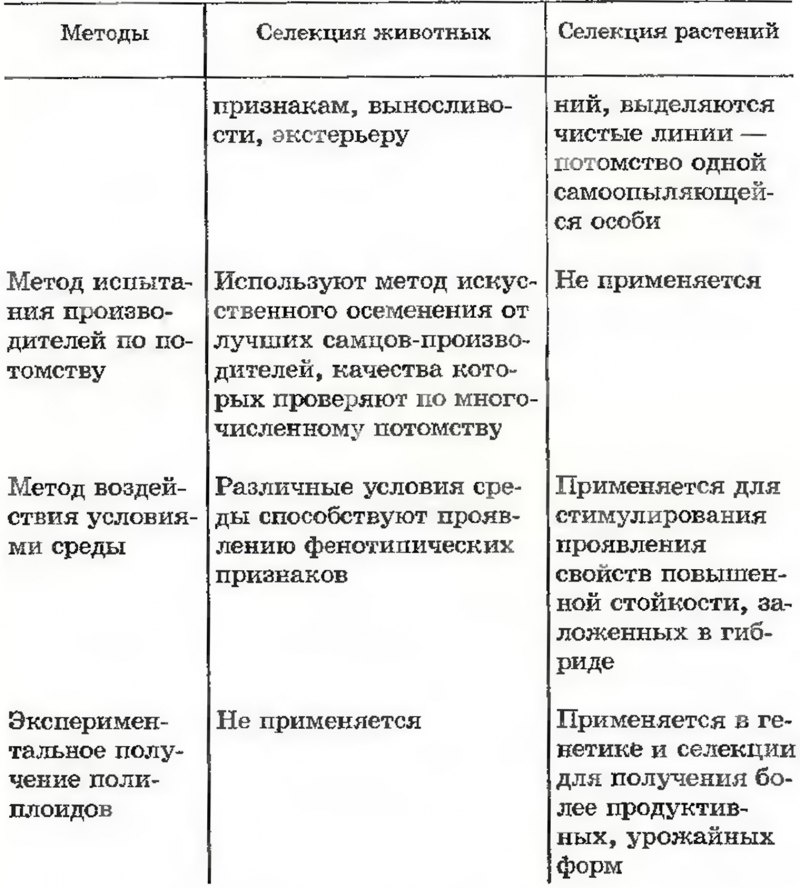
__________________
32. Генетика растений, методы и достижения.
Генетика растений, раздел генетики, изучающий наследственность и изменчивость высших растений (генетические исследования грибов и водорослей обычно относят к генетике микроорганизмов), Для генетического изучения растений, кроме методов, которыми пользуются в др. областях генетики (в частности гибридологического анализа), применяют следующие методы. С помощью моносомного анализа определяют роль каждой хромосомы в наследовании и развитии различных признаков растений. Этим методом (разработанным на дурмане) пользуются при изучении ряда аллополиплоидов (некоторых пшениц, хлопчатника), а также диплоидов (ячменя). Большое значение в Г. р. приобретает экспериментальный мутагенез который даёт огромное разнообразие новых форм, используемых в селекции, и ценный материал для изучения генетики отдельных видов растений. С помощью мутантов составляют генетические карты хромосом; на них исследуют действие изменённого гена (в гомо- и гетерозиготном состоянии) на развитие отдельных признаков в разных условиях среды, на физиологические и биохимические особенности растений. Изучение мутантов способствует выяснению эволюции того или иного вида. К методам исследования эволюции растений относятся также гибридизация и анализ конъюгации хромосома в мейозе у гибридов (неродственно хромосомы не конъюгируют). Важный метод — искусственный ресинтез существующих видов путём гибридизации и последующего удвоения числа хромосом (см. Полиплоидия). Значительную роль в эволюции растений, в том числе многих культурных (пшеницы, овса, хлопчатника, картофеля, плодовых и др.), играет аллополиплоидия. После открытия действия алкалоида колхицина, препятствующего расхождению удвоившихся хромосом к разным полюсам клетки, для получения новых, иногда очень ценных форм широко используется автополиплоидия. Сочетая методы отдалённой гибридизации и цитогенетики, изучают роль отдельных хромосом (и их участков) в наследовании признаков и разрабатывают приёмы, позволяющие получать вставки участков хромосом диких растений, обусловливающие развитие ценных признаков (например, устойчивости к ржавчине), в хромосомы культурных растений. Роль ядра и цитоплазмы в наследовании и развитии признаков исследуют, применяя отдалённую гибридизацию и анализируя природу мужской цитоплазматической стерильности, используемой при получении гетерозисных форм. В Г. р. широко исследуются апомиксис и явление самонесовместимости, т. е. неспособности растений к самооплодотворению, а также генетические особенности растений само- и перекрёстноопылителей, вегетативно и апомиктически размножающихся форм. В Г. р. всё больше проникают идеи и методы молекулярной биологии (гибридизация ДНК, ДНК — РНК, изучение изозимов и др.). Методы популяционной генетики и биометрии применяют в Г. р. для разграничения генотипических и паратипических элементов в общей фенотипической изменчивости признаков, что усиливает эффективность искусственного отбора. Все эти методы используют для улучшения хозяйственно ценных свойств с.-х. растений: урожайности, устойчивости к неблагоприятным условиям среды, ряда биохимических и технологических особенностей растения (или его зерна), особенностей развития (озимость, яровость, раннеспелость и т.д.). Из высших растений генетически наиболее изучены кукуруза, арабидопсис (растение семейства крестоцветных, «растительная дрозофила» — модельный объект генетических исследований), горох, томаты, ячмень. У этих растений методами гибридизации установлена локализация генов и составлены карты хромосом. Интенсивно изучается цитогенетика мягкой пшеницы — сложного 42-хромосомного аллополиплоида, возникшего в процессе эволюции при естественной гибридизации трёх разных злаков с последующим удвоением числа хромосом у гибридов. Вклад Г. р. в селекцию огромен. Это, например, использование гетерозиса в селекции кукурузы на основе мужской стерильности; введение высокоурожайным гибридам и сортам кормового ячменя генов, обеспечивающих высокое содержание лизина в зерне; создание низкорослых неполегающих высокоурожайных сортов пшеницы с использованием генов карликовости («зелёная революция» в Индии и др. странах); выведение урожайных и сахаристых триплоидных гибридов сахарной свёклы.
33. Эволюционные представления до Чарльза Дарвина.
Эволюционные представления до Чарльза Дарвина.
На Земле существуют не менее 2 млн.видов животных,до 0,5 млн.видов растений,сотни тысяч видов грибов и микроорганизмов.Как возникло великое многообразие видов и приспособленность их к среде обитания?Ответ дает научная теория эволюции живой природы,основы которой в 19 в.заложил великий английский ученый Чарлз Дарвин.
До Дарвина большинство биологов держалось представлений о постоянстве и неизменности живых организмов-видов столько,сколько их создал бог.Организмы и органыполностью соответствуют цели,которую якобы поставил творец.Сущность мировоззрения этого периода заключается в представлениях о постоянстве,неизменности и изначальной целесообразности природы.Такое мировоззрение получило название метафизического (греч."физис"-природа,"мета"-над).Метафизические представления потдерживались церквью и правящими кругами.
В 17-18 вв. накопилось множество описаний видов животных,растений,минералов.Огромную задачу систематизации этих материалов выполнил Карл Линней(1707-1778), шведский натуралист,врач.На основе сходства по одному-двум наиболее заметным признакам он классифицировал организмы на виды,роды,классы.Он правильно поместил в один отряд человека и человекообразных обезьян.Линней ввел в науку предложенный предшественниками двойные латинские названия-род и вид(Canis familiaris-собака домашняя).Латинские названия помогли общению ученых разных стран. Линней полностью разделял метафизические представления о природе,усматривая в ней изначальную целесообразность,якобы доказывающую "премудрость творца".Каждый вид он считал результатом отдельного творческого акта,неизменным и постоянным,не связанным с другими видами родством.К концу жизни,однако,под влиянием наблюдений в природе он признал,что иногда виды могут возникать путем скрещивания или в результате действий изменений среды.
Значение трудов Линнея огромно:он предложил систему животных и растений,лучшую из всех предыдущих;ввел двойные названия видов;усовершенствовал ботанический язык.
В начале 19 в.французский ученый Жан Батист Ламарк(1744-1829) изложил свои эволюционные идеи в труде "Философия зоологии".Лемарк подверг критеке идеи о постоянстве и неизменяемости видов.Он утверждал,что образование новых видов происходит очень медленно и потому незаметно.В процессе эволюции высшие формы жизни взяли начало от низших.
Значение трудов Ламарка для дальнейшего развития биологии огромно.Он первый изложил идеи эволюции живой природы,утверждавшие историческое развитие от простого к сложному.Он первым поставил вопрос о факторах-движущих силах эволюции. Тем не менее Ламарк ошибочно выводил факторы эволюции из будто бы присущего всему живому стремления к совершенству.Неверно объяснял причины возникновения приспособленности прямым влиянием условий окружающей среды.Неверно и утверждение об обязательном появлении только полезных изменений и их наследовании. Итак наука 18-начала 19 в.не могла правильно объяснить движущие силы развития органического мира.Перед нею встали вопросы:как возникло огромное многообразие видов?Как объяснить приспособленность организмов к условиям окружающей среды? Почему в процессе эволюции происходит повышение организации живых существ?
Возникновение учения Чарльза Дарвина.
Возникновение учения Ч.Дарвина способствовали общественно-экономические предпосылки.В первой половине 19 в. в странах Западной Европы,особенно в Англии,интенсивно развивался капитализм,который дал импульс развитию науки,промышленности ,техники.Спрос промышленности на сырье и населения растущих городов на продукты питания способствовал развитию сельского хозяйства.
Другая предпосылка появления дарвинизма-успехи естественных наук.Описания систематических групп живых организмов приводили к мысли о возможности их родства. У многих животных сравнением установили единый план в строении тела и органов. Исследования ранних стадий развития зародышей хордовых выявили их поразительное сходство.Изучение ископаемых растений и животных раскрыло последовательную смену низкоорганизованных форм жизни более высокоорганизованными.
Обширные материалы заморских экспидиций,выведение новых пород животных и сортов растений не согласовывались с метафизическим мировоззрением.Нужен был гениальный ум,который сумел бы обобщить огромный материал в свете определенной идеи, связать стройной системой рассуждений.Таким ученым оказался Чарлз Дарвин (1809-1882).
С детства Ч.Дарвин увлекался сбором коллекций,химическими опытами,наблюдениями за животными.Студентом изучал научную литературу,овладел методикой полевых исследований.Ч.Дарвин на корабле "Бигл"(англ.-ищейка) совершил кругосветное путешествие.Он исследовал геологическое строение,флору и фауну многих стран,отправил в Англию огромное количество коллекций.
В Южной Америке,сравнив найденные останки вымерших животных с современными, Ч.Дарвин предположил их родство.На Галапагосских островах он нашел нигде более не встречающиеся виды ящериц,черепах,птиц.Они близки к южноамериканским. Галапагосские острова вулканического происхождения,и потому Ч.Дарвин предположил,что на них виды попали с материка и постепенно изменились.В Австралии его заинтересовали сумчатые и яйцекладущие,которые вымерли в других местах земного шара.Австралия как материк обособилась,когда еще не возникли высшие млекопитающие.Сумчатые и яйцекладущие развивались здесь независимо от эволюции млекопитающих на других материках.Так постепенно крепло убеждение в изменяемости видов и происхождении одних от других.Первые записио происхождении видов Дарвин сделал во время кругосветногопутешествия.
Основные положения учения Дарвина.
После путешествия Дарвин в течение 20 лет упорно работал над созданием эволюционного учения и опубликовал его в труде "Происхождение видов путем естественного отбора,или Сохранение благоприятствуемых пород в борьбе за жизнь"(1859).В дальнейших произведениях Дарвин развивал и углублял различные стороны основной проблемы-происхождение видов.В книге "Изменение домашних животных и культурных растений" на огромном фактическом материале он показал закономерности эволюции пород домашних животных и сортов культурных растений.
В труде "Происхождение человека и половой отбор" Дарвин применил эволюционную теорию для объяснения происхождения человека от животных.Дарвину принадлежат капитальные труды по ботанике,зоологии и геологии,в которых детально разработанны отдельные вопросы эволюционной теории.
Главная заслуга Дарвина в том,что он раскрыл движущие силы эволюции.Он материалистически объяснил возникновение и относительный характер приспособленности действием только естественных законов,без вмешательства сверхестественных сил. Учение Дарвина в корне подрывало метафизические представления о постоянстве видов и сотворении их богом.Каковы же движущие силы эволюции пород домашних животных,сортов культурных растений и видов в дикой природе?