
Рис 3.4. Цитологические основы расщепления признаков при.дигиб-ридном скрещивании.








Гомозиготные родители (ААВВ и aabb) формируют только один тип гамет с доминантными (АВ) или с рецессивными (ab) аллелями. При слиянии таких гамет образуется единообразное первое поколение гибридов — гибрид дигетерозиготен (АаВb), но так как у него присутствуют гены А и B, то по фенотипу он сходен с одним из родителей.
В тех случаях, когда необходимо указать, что те или иные гены находятся в гомологичных хромосомах, в генетических формулах зигот хромосомы принято изображать в виде двух черточек или одной с указанием обоих аллелей гена. Формула дигетерозиготы может быть записана так: 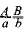 . Поскольку гаметы содержат только по одной из гомологичных хромосом и соответственно по одному аллелю каждого гена, то их формулы могут быть записаны так:
. Поскольку гаметы содержат только по одной из гомологичных хромосом и соответственно по одному аллелю каждого гена, то их формулы могут быть записаны так: 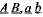 и т. д.
и т. д.
В дальнейшем у гибридных организмов ло причине случайности расхождения отцовских и материнских хромосом каждой пары в процессе мейоза ген А может попасть в одну гамету с геном В или с геном Ь. Точно так же ген а может оказаться в одной гамете с геном В или с геном b. Поэтому гибриды образуют четыре типа гамет: 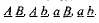 Образование всех четырех типов гамет равновероятно, т. е. все они образуются в равных количествах. Свободное сочетание таких гамет в процессах оплодотворения заканчивается образованием 16 типов зигот, а значит, и потомков (см. рис. 3.4).
Образование всех четырех типов гамет равновероятно, т. е. все они образуются в равных количествах. Свободное сочетание таких гамет в процессах оплодотворения заканчивается образованием 16 типов зигот, а значит, и потомков (см. рис. 3.4).
Оли распадаются на четыре фенотипических класса: доминантные по обоим признакам — 9 частей, доминантные по первому и рецессивные по второму признаку — 3 части, рецессивные по первому и доминантные по второму — 3 части, рецессивные по обоим признакам — 1 часть. Генотипических классов 9: 1AABB, 2ААВb, 1AAbb, 1Aabb, 4AaBb, 2AaBB, 1aaBB, 2aaBb, 1aabb.
Полигибридное скрещивание. Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании, т. е. когда родители различаются по аллелям трех и более генов, а в F1 образуются три- и по дигетерозиготы. Соотношение генотипических и фенотипических классов в F2 три- и полигибридных скрещиваний, а также число типов гамет (и число фенотипов) у гибридов F1 определяются простыми формулами: при
моногибридном скрещивании число типов гамет равно 2, при дигибридном 4(22), а при полигибридном — 2n; число генотипов равно соответственно 3,9(32) и 3n.
Опираясь на независимость наследования разных пар аллелей, можно также любые сложные расщепления представить как произведение от соответствующего числа независимых моногиб-ридиых скрещивании. Общая формула определения фенотипических классов при полигибридном скрещивании имеет вид (3:1)n, где п равно числу пар признаков, по которым идет расщепление. Для моногибрида эта формула соответственно имеет вид (3:1); дигибрида — 9:3:3:1 или(3:1)2;тригибрида — (3:1)3. Расщепление по генотипу имеет вид (1:2; 1)n, где п — число расщепляющихся пар аллелей.
Известно, что каждый организм гетерозиготен по многим генам. Если предположить, что человек, у которого отдельные пары хромосом содержат не одну, а сотни пар аллелей, гетерозиготен хотя бы по 20 генам, то число типов гамет у такой полигетерозиго-ты составит 220 = 1 048 576. Эта цифра дает определенное представление о потенциальных возможностях комбинативной изменчивости. Поэтому каждый человек обладает неповторимой индивидуальностью. На Земле нет двух людей, совершенно одинаковых по наследственности, за исключением однояйцевых близнецов.
Таким образом, третий закон Менделя (закон независимого наследования признаков) еще раз демонстрирует дискретный характер генетического материала. Это проявляется в независимом комбинировании аллелей разных генов и в их независимом дей-ствии — фенотипическом выражении.
Дискретность гена определяется тем, что он контролирует присутствие или отсутствие отдельной биохимической реакции, от которой зависит развитие или подавление определенного признака организма. Очевидно, если несколько генов определяют какое-либо одно свойство или один признак (форма гребня у кур, окраска глаз у дрозофилы, длина колоса у пшеницы и т. д.), они должны взаимодействовать между собой. Отсюда следует, что понятие «наследование признаков» употребляется, скорее всего, как образное выражение, поскольку в действительности наследуются не сами признаки, а гены. Признаки формируются в ходе индивидуального развития организма, обусловливаются генотипом и влиянием внешней среды.
1ААВВ, lAABb, 1AAbb, lAabb, AAaBb, 2AaBB, 1aaBB,
МЕНДЕЛЯ ЗАКОНЫ, основные закономерности наследования, открытые Г. Менделем. В 1856–1863 гг. Мендель провёл обширные, тщательно спланированные опыты по гибридизации растений гороха. Для скрещиваний он отбирал константные сорта (чистые линии), каждый из которых при самоопылении устойчиво воспроизводил в поколениях одни и те же признаки. Сорта различались альтернативными (взаимоисключающими) вариантами какого-либо признака, контролируемого парой аллельных генов (аллелей). Напр., окраской (жёлтая или зелёная) и формой (гладкая или морщинистая) семян, длиной стебля (длинный или короткий) и т. д. Для анализа результатов скрещиваний Мендель применил математические методы, что позволило ему обнаружить ряд закономерностей в распределении родительских признаков у потомков. Традиционно в генетике принимают три закона Менделя, хотя сам он формулировал лишь закон независимого комбинирования. Первый закон, или закон единообразия гибридов первого поколения, утверждает, что при скрещивании организмов, различающихся аллельными признаками, в первом поколении гибридов проявляется лишь один из них – доминантный, а альтернативный ему, рецессивный, остаётся скрытым (см. Доминантность, Рецессивность). Напр., при скрещивании гомозиготных (чистых) сортов гороха с жёлтой и зелёной окраской семян у всех гибридов первого поколения окраска была жёлтой. Значит, жёлтая окраска – доминантный признак, а зелёная – рецессивный. Первоначально этот закон называли законом доминирования. Вскоре было обнаружено его нарушение – промежуточное проявление обоих признаков, или неполное доминирование, при котором, однако, сохраняется единообразие гибридов. Поэтому современное название закона более точное.
Второй закон, или закон расщепления, гласит, что при скрещивании между собой двух гибридов первого поколения (или при их самоопылении) во втором поколении проявляются в определённом соотношении оба признака исходных родительских форм. В случае жёлтой и зелёной окраски семян их соотношение было 3:1, т. е. расщепление по фенотипу происходит так, что у 75 % растений окраска семян доминантная жёлтая, у 25 % – рецессивная зелёная. В основе такого расщепления лежит образование гетерозиготными гибридами первого поколения в равном отношении гаплоидных гамет с доминантными и рецессивными аллелями. При слиянии гамет у гибридов 2-го поколения образуется 4 генотипа – два гомозиготных, несущих только доминантные и только рецессивные аллели, и два гетерозиготных, как у гибридов 1-го поколения. Поэтому расщепление по генотипу 1:2:1 даёт расщепление по фенотипу 3:1 (жёлтую окраску обеспечивает одна доминантная гомозигота и две гетерозиготы, зелёную – одна рецессивная гомозигота).
Третий закон, или закон независимого комбинирования, утверждает, что при скрещивании гомозиготных особей, отличающихся по двум и более парам альтернативных признаков, каждая из таких пар (и пар аллельных генов) ведёт себя независимо от других пар, т. е. и гены, и соответствующие им признаки наследуются в потомстве независимо и свободно комбинируются во всех возможных сочетаниях. Он основан на законе расщепления и выполняется в том случае, если пары аллельных генов расположены в разных гомологичных хромосомах.
Часто как один из законов Менделя приводится и закон чистоты гамет, утверждающий, что в каждую половую клетку попадает только один аллельный ген. Но этот закон был сформулирован не Менделем.
Непонятый современниками, Мендель обнаружил дискретную («корпускулярную») природу наследственности и показал ошибочность представлений о «слитной» наследственности. После переоткрытия забытых законов основанное на экспериментах учение Менделя получило название менделизм. Его справедливость была подтверждена хромосомной теорией наследственности.
Третий закон Менделя – закон независимого комбинирования: «При скрещивании двух гомозиготных особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях». Третий закон Менделя При дигибридном скрещивании двух дигетерозигот (особей F1) между собой, во втором поколении гибридов (F2) будет наблюдаться расщепление признаков по фенотипу в соотношении 9:3:8:1, т. е. сочетание двух вариантов обоих признаков позволит получить четыре группы фенотипов в потомстве. Если же рассмотреть наследование каждого признака в отдельности, то по каждому из них будет наблюдаться расщепление 8:1.
24. Сцепление генов, закон Томаса Моргана.
Закон Томаса Моргана
Дальнейшие исследования генетиков показали, что законы Менделя о независимом наследовании признаков при дигибридном скрещивании применимы лишь тогда, когда разные гены располагаются в разных парах гомологичных хромосом. В том случае,если два гена находятся в одной паре гомологичных хромосом, расщепление в потомстве гибридов будет другим.
У любого организма генов значительно больше, чем хромосом. Например, у человека имеется около миллиона генов, а хромосом всего 23 пары. Следовательно,в одной хромосоме размещается в среднем несколько тысяч генов. Гены,расположенные в одной хромосоме, называют сцепленными. Все гены этой хромосомы образуют группу сцепления, которая при мейозе обычно попадает в одну гамету.
Значит, гены, входящие в одну группу сцепления, не подчиняются закону независимого наследования, а при дигибридном скрещивании вместо ожидаемого расщепления по фенотипу в соотношении 9:3:3:1 дают соотношение 3:1, как при моногибридном скрещивании.
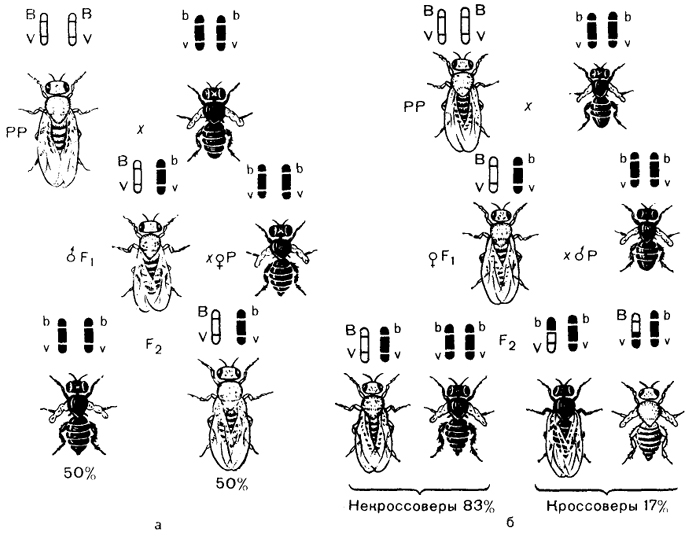
Закономерности сцепленного наследования были установлены американским биологом Томасом Морганом (1866-1945). В качестве объекта он использовал плодовую муху дрозофилу. У дрозофилы окраску тела и длину крыльев определяют следующие пары аллелей: А - серое тело, а - черное тело, В - длинные крылья, b- зачаточные крылья. Гены, отвечающие за окраску тела и длину крыльев,находятся в одной паре гомологичных хромосом и наследуются сцепленно.
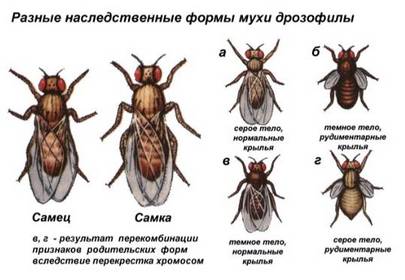
При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой,имеющей черное тело и зачаточные крылья, все гибриды первого поколения имели серую окраску тела и длинные крылья.
При дальнейшем скрещивании между собой гибридных мух первого поколения в F2не произошло ожидаемого расщепления по фенотипу 9:3:3:1. Вместо этого в F2были получены мухи с родительскими фенотипами в соотношении примерно 3:1.Появление в F2 двух фенотипов вместо четырех позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме. Так был установлен закон Т.Моргана: гены, расположенные в одной хромосоме, наследуются совместно - сцепленно, то есть наследуются преимущественно вместе.
Однако при дигибридном скрещивании при сцепленном наследовании признаков не всегда появляются особи только двух фенотипов. Иногда появляются особи еще двух фенотипов с перекомбинацией (новым сочетанием) родительских признаков: серое тело - зачаточные крылья, черное тело - длинные крылья. (Особей с такими фенотипами немного - около 8,5% каждого типа.) Почему же нарушается сцепление генов и появляются особи с новыми фенотипами? Было установлено, что сцепление генов может быть полным и неполным.
Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с черным телом и зачаточными крыльями. Расщепление по фенотипу в этом случае будет 1:1, то есть наблюдается полное сцепление генов в одной хромосоме.
При скрещивании серой длиннокрылой самки с самцом, имеющим черное тело и зачаточные крылья, расщепление по фенотипу будет примерно 41,5:41,5:8,5:8,5,что характеризует неполное сцепление. Причина нарушения сцепления заключается в том, что в ходе мейоза происходит кроссинговер и гомологичные хромосомы обмениваются своими участками. В результате гены, расположенные в одной из гомологичных хромосом, оказываются в другой хромосоме. Возникают новые сочетания признаков.
У самцов дрозофил в мейозе кроссинговер не происходит, поэтому при скрещивании серого длиннокрылого самца дрозофилы с рецессивной самкой с черным телом и зачаточными крыльями сцепление будет полным. Неполное сцепление наблюдается в том случае, если самка гетерозиготна, а самец гомозиготен. В данном примере кроссинговер происходит примерно у 17% самок.
Таким образом, если не происходит перекреста хромосом и обмена генами, то наблюдается полное сцепление генов. При наличии кроссинговера сцепление генов бывает неполным. Благодаря перекресту хромосом возникают новые сочетания генов и признаков. Чем дальше друг от друга расположены гены в хромосоме, тем больше вероятность перекреста между ними и обмена участками хромосом.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах: единице расстояния между генами, находящимися в одной хромосоме,соответствует 1% кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид.
Сравните результаты скрещивание дрозофил:
а) Полное сцепление без кроссинговера
б) С частотой кроссинговера равной 17 %
Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности:
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3. Гены расположены в хромосомах в определенной линейной последовательности;
4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом;
6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);