
Теории стационарного состояния и панспермии








Обе эти теории представляют собой взаимодополняющие элементы единой картины мира, сущность которой заключается в следующем: вселенная существует вечно и в ней вечно существует жизнь (стационарное состояние). Жизнь переносится с планеты на планету путешествующими в космическом пространстве «семенами жизни», которые могут входить в состав комет и метеоритов (панспермия). Подобных взглядов на происхождение жизни придерживался, в частности, основоположник учения о биосфере академик В.И. Вернадский.
Однако теория стационарного состояния, предполагающая бесконечно долгое существование вселенной, не согласуется с данными современной астрофизики, согласно которым вселенная возникла сравнительно недавно (около 16 млрд лет т.н.) путем первичного взрыва.
Очевидно, что обе теории (панспермии и стационарного состояния) вообще не предлагают объяснения механизма первичного возникновения жизни, перенося его на другие планеты (панспермия) либо отодвигая по времени в бесконечность (теория стационарного состояния).
Теория биохимической эволюции (теория А.И. Опарина)
Из всех теорий происхождения жизни наиболее распространенной и признанной в научном мире является теория биохимической эволюции, предложенная в 1924 г. советским биохимиком академиком А.И. Опариным (в 1936 г. он подробно изложил ее в своей книге «Возникновение жизни»).
Сущность этой теории состоит в том, что биологической эволюции — т.е. появлению, развитию и усложнению различных форм живых организмов, предшествовала химическая эволюция — длительный период в истории Земли, связанный с появлением, усложнением и совершенствованием взаимодействия между элементарными единицами, «кирпичиками», из которых состоит все живое — органическими молекулами.
Предбиологическая (химическая) эволюция
По мнению большинства ученых (в первую очередь астрономов и геологов), Земля сформировалась как небесное тело около 5 млрд лет т.н. путем конденсации частиц вращавшегося вокруг Солнца газопылевого облака.
Под влиянием сил сжатия частицы, из которых формируется Земля, выделяют огромное количество тепла. В недрах Земли начинаются термоядерные реакции. В результате Земля сильно разогревается. Таким образом, 5 млрд лет т.н. Земля представляла собой несущийся в космическом пространстве раскаленный шар, температура поверхности которою достигала 4000-8000°С (рис. 2.4.1.1).
Постепенно, за счет излучения тепловой энергии в космическое пространство, Земля начинает остывать. Около 4 млрд лет т.н. Земля остывает настолько, что на ее поверхности формируется твердая кора; одновременно из ее недр вырываются легкие, газообразные вещества, поднимающиеся вверх и формирующие первичную атмосферу. По составу первичная атмосфера существенно отличалась от современной. Свободный кислород в атмосфере древней Земли, по-видимому, отсутствовал, а в ее состав входили вещества в восстановленном состоянии, такие, как водород (Н2), метан (СН4), аммиак (NH3), пары воды (Н2О), а возможно, также азот (N2), окись и двуокись углерода (СО и С02).
Восстановительный характер первичной атмосферы Земли чрезвычайно важен для зарождения жизни, поскольку вещества в восстановленном состоянии обладают высокой реакционной способностью и в определенных условиях способны взаимодействовать друг с другом, образуя органические молекулы. Отсутствие в атмосфере первичной Земли свободного кислорода (практически весь кислород Земли был связан в виде окислов) также является важной предпосылкой возникновения жизни, поскольку кислород легко окисляет и тем самым разрушает органические соединения. Поэтому при наличии в атмосфере свободного кислорода накопление на древней Земле значительного количества органических веществ было бы невозможно.
Около 5 млрд лет т.п. — возникновение Земли как небесного тела; температура поверхности — 4000-8000°С
Около 4 млрд лет т.н. - формирование земной коры и первичной атмосферы
При температуре 1000°С — в первичной атмосфере начинается синтез простых органических молекул
Энергию для синтеза дают:

Температура первичной атмосферы ниже 100°С — формирование первичного океана -
Синтез сложных органических молекул — биополимеров из простых органических молекул:
§ простые органические молекулы — мономеры
§ сложные органические молекулы — биополимеры
Рис. 2.4.1.1. Основные этапы химической эволюции
Когда температура первичной атмосферы достигает 1000°С, в ней начинается синтез простых органических молекул, таких, как аминокислоты, нуклеотиды, жирные кислоты, простые сахара, многоатомные спирты, органические кислоты и др. Энергию для синтеза поставляют грозовые разряды, вулканическая деятельность, жесткое космическое излучение и, наконец, ультрафиолетовое излучение Солнца, от которого Земля еще не защищена озоновым экраном, причем именно ультрафиолетовое излучение ученые считают основным источником энергии для абиогенного (т.е. проходящею без участия живых организмов) синтеза органических веществ.
Признанию и широкому распространению теории А.И. Опарина во многом способствовало то, что процессы абиогенного синтеза органических молекул легко воспроизводятся в модельных экспериментах.
Возможность синтеза органических веществ из неорганических была известна с начала 19 в. Уже в 1828 г. выдающийся немецкий химик Ф. Вёлер синтезировал органическое вещество — мочевину из неорганическою — циановокислого аммония. Однако возможность абиогенного синтеза органических веществ в условиях, близких к условиям древней Земли, была впервые показана в опыте С. Миллера.
В 1953 г. молодой американский исследователь, студент- дипломник Чикагского университета Стенли Миллер воспроизвел в стеклянной колбе с впаянными в нес электродами первичную атмосферу Земли, которая, по мнению ученых того времени, состояла из водорода метана СН4, аммиака NH, и паров воды Н20 (рис. 2.4.1.2). Через эту газовую смесь С. Миллер в течение недели пропускал электрические разряды, имитирующие грозовые. По окончании эксперимента в колбе были обнаружены α-аминокислоты (глицин, аланин, аспарагин, глутамин), органические кислоты (янтарная, молочная, уксусная, гликоколовая), у-оксимасляная кислота и мочевина. При повторении опыта С. Миллеру удалось получить отдельные нуклеотиды и короткие полинуклеотидные цепочки из пяти-шести звеньев.
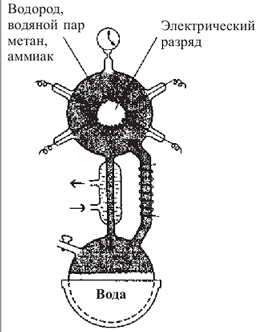
Рис. 2.4.1.2. Установка С. Миллера
В дальнейших опытах по абиогенному синтезу, проводимых различными исследователями, использовались не только электрические разряды, но и другие виды энергии, характерные для древней Земли, — космическое, ультрафиолетовое и радиоактивное излучения, высокие температуры, присущие вулканической деятельности, а также разнообразные варианты газовых смеси, имитирующих первичную атмосферу. В результате был получен практически весь спектр органических молекул, характерных для живого: аминокислоты, нуклеотиды, жироподобные вещества, простые сахара, органические кислоты.
Более того, абиогенный синтез органических молекул может происходить на Земле и в настоящее время (например, в процессе вулканической деятельности). При этом в вулканических выбросах можно обнаружить не только синильную кислоту HCN, являющуюся предшественником аминокислот и нуклеотидов, но и отдельные аминокислоты, нуклеотиды и даже такие сложные по строению органические вещества, как порфирины. Абиогенный синтез органических веществ возможен не только на Земле, но и в космическом пространстве. Простейшие аминокислоты обнаружены в составе метеоритов и комет.
Когда температура первичной атмосферы опустилась ниже 100°С, на Землю обрушились горячие дожди и появился первичный океан. С потоками дождя в первичный океан поступали абиогенно синтезированные органические вещества, что превратило его, но образному выражению английского биохимика Джона Холдейна, в разбавленный «первичный бульон». По-видимому, именно в первичном океане начинаются процессы образования из простых органических молекул — мономеров сложных органических молекул — биополимеров (см. рис. 2.4.1.1).
Однако процессы полимеризации отдельных нуклеогидов, аминокислот и Сахаров — это реакции конденсации, они протекают с отщеплением воды, следовательно, водная среда способствует не полимеризации, а, напротив, гидролизу биополимеров (т.е. разрушению их с присоединением воды).
Образование биополимеров (в частности, белков из аминокислот) могло происходить в атмосфере при температуре около 180°С, откуда они смывались в первичный океан с атмосферными осадками. Кроме того, возможно, на древней Земле аминокислоты концентрировались в пересыхающих водоемах и полимеризовались в сухом виде под действием ультрафиолетового света и тепла лавовых потоков.
Несмотря на то что вода способствует гидролизу биополимеров, в живой клетке синтез биополимеров осуществляется именно в водной среде. Этот процесс катализируют особые белки-катализаторы — ферменты, а необходимая для синтеза энергия выделяется при распаде аденозинтрифосфорной кислоты — АТФ. Возможно, синтез биополимеров в водной среде первичного океана катализировался поверхностью некоторых минералов. Экспериментально показано, что раствор аминокислоты аланина может полимеризоваться в водной среде в присутствии особого вида глинозема. При этом образуется пептид полиаланин. Реакция полимеризации аланина сопровождается распадом АТФ.
Полимеризация нуклеотидов проходит легче, чем полимеризация аминокислот. Показано, что в растворах с высокой концентрацией солей отдельные нуклеотиды самопроизвольно полимеризуются, превращаясь в нуклеиновые кислоты.
Жизнь всех современных живых существ — это процесс непрерывного взаимодействия важнейших биополимеров живой клетки — белков и нуклеиновых кислот.
Белки — это «молекулы-рабочие», «молекулы-инженеры» живой клетки. Характеризуя их роль в обмене веществ, биохимики часто используют такие образные выражения, как «белок работает», «фермент ведет реакцию». Важнейшая функция белков- каталитическая. Как известно, катализаторы — это вещества, которые ускоряют химические реакции, но сами в конечные продукты реакции не входят. Бачки-катализаторы называются ферментами. Ферменты в согни и тысячи раз ускоряют реакции обмена веществ. Обмен веществ, а значит, и жизнь без них невозможны.
Нуклеиновые кислоты — это «молекулы-компьютеры», молекулы — хранители наследственной информации. Нуклеиновые кислоты хранят информацию не обо всех веществах живой клетки, а только о белках. Достаточно воспроизвести в дочерней клетке белки, свойственные материнской клетке, чтобы они точно воссоздали все химические и структурные особенности материнской клетки, а также свойственный ей характер и темпы обмена веществ. Сами нуклеиновые кислоты также воспроизводятся благодаря каталитической активности белков.
Таким образом, тайна зарождения жизни — это тайна возникновения механизма взаимодействия белков и нуклеиновых кислот. Какими же сведениями об этом процессе располагает современная наука? Какие молекулы явились первичной основой жизни — белки или нуклеиновые кислоты?
Ученые полагают, что несмотря на ключевую роль белков в обмене веществ современных живых организмов, первыми «живыми» молекулами были не белки, а нуклеиновые кислоты, а именно рибонуклеиновые кислоты (РНК).
В 1982 г. американский биохимик Томас Чек открыл автокаталитические свойства РНК. Он экспериментально показал, что в среде, содержащей в высокой концентрации минеральные соли, рибонуклеотиды спонтанно (самопроизвольно) полимеризуются, образуя полинуклеотиды — молекулы РНК. На исходных поли- нуклеотидных цепях РНК, как на матрице, путем спаривания комплементарных азотистых оснований образуются РНК-ко- пии. Реакция матричного копирования РНК катализируется исходной молекулой РНК и не требует участия ферментов либо других белков.
Дальнейшие события достаточно хорошо объясняются процессом, который можно было бы назвать «естественным отбором» на уровне молекул. При самокопировании (самосборке) молекул РНК неизбежно возникают неточности, ошибки. Содержащие ошибки копии РНК снова копируются. При повторном копировании вновь могут возникнуть ошибки. В результате популяция молекул РНК на определенном участке первичного океана будет неоднородна.
Поскольку параллельно с процессами синтеза идут и процессы распада РНК, в реакционной среде будут накапливаться молекулы, обладающие либо большей стабильностью, либо лучшими автокаталитическими свойствами (т.е. молекулы, которые быстрее себя копируют, быстрее «размножаются»).
На некоторых молекулах РНК, как на матрице, может происходить самосборка небольших белковых фрагментов — пептидов. Вокруг молекулы РНК образуется белковый «чехол».
Наряду с автокаталитическими функциями Томас Чек обнаружил у молекул РНК и явление самосплайсинга. В результате самосплайсинга участки РНК, не защищенные пептидами, самопроизвольно удаляются из РНК (они как бы «вырезаются» и «выбрасываются»), а оставшиеся участки РНК, кодирующие белковые фрагменты, «срастаются», т.е. самопроизвольно объединяются в единую молекулу. Эта новая молекула РНК уже будет кодировать большой сложный белок (рис. 2.4.1.3).
По-видимому, первоначально белковые чехлы выполняли в первую очередь, защитную функцию, предохраняя РНК от разрушения и повышая тем самым ее стабильность в растворе (такова функция белковых чехлов и у простейших современных вирусов).
Очевидно, что на определенном этапе биохимической эволюции преимущество получили молекулы РНК, кодирующие не только защитные белки, но и белки-катализаторы (ферменты), резко ускоряющие скорость копирования РНК. По-видимому, именно таким образом и возник процесс взаимодействия белков и нуклеиновых кислот, который мы в настоящее время называем жизнью.
В процессе дальнейшего развития, благодаря появлению белка с функциями фермента — обратной транскриптазы, на одно- цепочечных молекулах РНК стали синтезироваться состоящие из двух цепей молекулы дезоксирибонуклеиновой кислоты (ДНК). Отсутствие у дезоксирибозы ОН-группы в 2' положении делает молекулы ДНК более стабильными по отношению к гидролитическому расщеплению в слабощелочных растворах, а именно слабощелочной была реакция среды в первичных водоемах (эта реакция среды сохранилась и в цитоплазме современных клеток).
Где же происходило развитие сложного процесса взаимодействия белков и нуклеиновых кислот? По теории А.И. Опарина, местом зарождения жизни стали так называемые коацерватные капли.
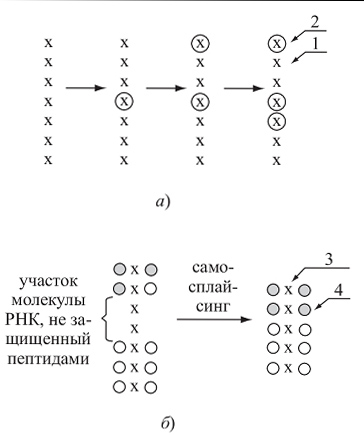
Рис. 2.4.1.3.
Гипотеза возникновения взаимодействия белков и нуклеиновых кислот:
а) в процессе самокопирования РНК накапливаются ошибки (1 — нуклеотиды, соответствующие исходной РНК; 2 — нуклеотиды, не соответствующие исходной РНК, — ошибки в копировании); б) на часть молекулы РНК за счет ее физико-химических свойств «налипают» аминокислоты (3 — молекула РНК; 4 — аминокислоты), которые, взаимодействуя друг с другом, превращаются в короткие белковые молекулы — пептиды.
В результате свойственного молекулам РНК самосплайсинга незащищенные пептидами участки молекулы РНК разрушаются, а оставшиеся «срастаются» в единую молекулу, кодирующую крупный белок.
В результате возникает молекула РНК, покрытая белковым чехлом (сходное строение имеют и наиболее примитивные современные вирусы, например вирус табачной мозаики)
Явление коацервации состоит в том, что в некоторых условиях (например, в присутствии электролитов) высокомолекулярные вещества отделяются от раствора, но не в форме осадка, а в виде более кон центрирован но го раствора — коацервата. При встряхивании коацерват распадается на отдельные мелкие капельки. В воде такие капли покрываются стабилизирующей их гидратной оболочкой (оболочкой из молекул воды) — рис. 2.4.1.4.
Коацерватные капли обладают некоторым подобием обмена веществ: иод воздействием чисто физико-химических сил они могут избирательно впитывать из раствора некоторые вещества и выделять в окружающую среду продукты их распада. За счет избирательного концентрирования веществ из окружающей среды они могут расти, а но достижении определенного размера начинают «размножаться», отпочковывая маленькие капельки, которые, в свою очередь, могут расти и «почковаться».
Возникшие в результате концентрирования белковых растворов коацерватные капли в процессе перемешивания под действием волн и ветра могут покрываться оболочкой из липи- дов: одинарной, напоминающей мицеллы мыла (при однократном отрыве капли от поверхности воды, покрытой липидным слоем), либо двойной, напоминающей клеточную мембрану (при повторном падении капли, покрытой однослойной липидной мембраной, на липидную пленку, покрывающую поверхность водоема — рис. 2.4.1.4).
Процессы возникновения коацерватных капель, их роста и «почкования», а также «одевания» их мембраной из двойного липидного слоя легко моделируются в лабораторных условиях.
Для коацерватных капель также существует процесс «естественного отбора», при котором в растворе сохраняются наиболее стабильные капли.
Несмотря на внешнее сходство коацерватных капель с живыми клетками, у коацерватных капель отсутствует главный признак живого — способность к точному самовоспроизведению, самокопированию. Очевидно, предшественниками живых клеток явились такие коацерватные капли, в состав которых вошли комплексы молекул-репликаторов (РНК или ДНК) и кодируемых ими белков. Возможно, комплексы РНК-белок длительное время существовали вне коацерватных капель в виде так называемого «свободноживущего гена», а возможно, их формирование проходило непосредственно внутри некоторых коацерватных капель.
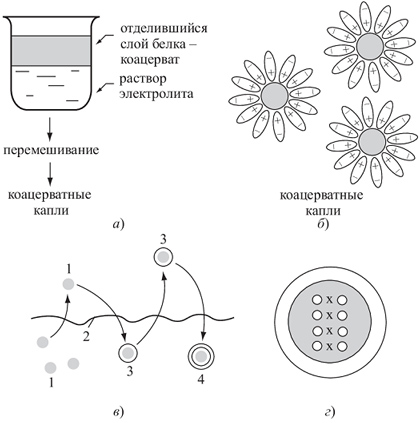
Рис 2.4.1.4.
Возможный путь перехода от коацерватных капель к примитивным клешам:
а) образование коацсрвата; 6) стабилизация коацерватных капель в водном растворе; в) — формирование вокруг капли двойного липидного слоя, похожего на клеточную мембрану: 1 — коацерватная капля; 2 — мономолекулярный слой липида на поверхности водоема; 3 — формирование вокруг капли одинарного липидного слоя; 4 — формирование вокруг капли двойного липидного слоя, похожего на клеточную мембрану; г) — коацерватная капля, окруженная двойным липидным слоем, с вошедшим в ее состав белково-нуклеотидным комплексом — прообраз первой живой клетки
Исключительно сложный, не до конца понятный современной науке процесс возникновения жизни на Земле прошел с исторической точки зрения чрезвычайно быстро. Уже 3,5 млрд лет т.н. химическая эволюция завершилась появлением первых живых клеток и началась биологическая эволюция.
Эры развития жизни на Земле
| ЭРЫ | Периоды и их продолжительность (млн. лет) | Животный и растительный мир | |
| Назв. и продолжит. | Возраст | ||
| Кайнозойская (новой жизни), 67 | 67 | Антропоген, 1,5 | Появление и развитие человека. Животный и растительный мир принял современный облик. |
| Неоген, 23,5 | Господство млекопитающих, птиц. | ||
| Палеоген, 42 | Появление хвостатых лемуров, долгопятов, позднее - парапитеков, дриопитеков. Бурный расцвет насекомых. Продолжается вымирание крупных пресмыкающихся. Исчезают многие группы головоногих моллюсков. Господство покрытосеменных растений. | ||
| Мезозойская (средней жизни), 163 | 230 | Меловой, 70 | Появление высших млекопитающих и настоящих птиц, хотя и зубастые птицы еще не распространены. Преобладают костистые рыбы. Сокращение папоротников и голосеменных. Появление и распространение покрытосеменных. |
| Юрский, 58 | Господство пресмыкающихся. Появление археоптерикса. Процветание головоногих моллюсков. Господство голосеменных растений. | ||
| Триасовый, 35 | Начало расцвета пресмыкающихся. Появление первых млекопитающих, настоящих костистых рыб. | ||
| Палеозойская (древней жизни), 340 | Возможно, 570 | Пермский, 55 | Быстрое развитие пресмыкающихся. Возникновение зверозубых пресмыкающихся. Вымирание трилобитов. Исчезновение каменноугольных лесов. Богатая флора голосеменных. |
| Каменноугольный, 75-65 | Расцвет земноводных. Возникновение первых пресмыкающихся. Появление летающих форм насекомых, пауков, скорпионов. Заметное уменьшение трилобитов. Расцвет папоротникообразных. Появление семенных папоротников. | ||
| Девонский, 60 | Расцвет щитковых. Появление кистеперых рыб. Появление стегоцефалов. Распространение на суше споровых растений. | ||
| Силурийский, 30 | Пышное развитие кораллов, трилобитов. Появление бесчелюстных позвоночных - щитковых. Выход растений на сушу - псилофиты. Широкое распространение водорослей. | ||
| Ордовийский, 60 Кембрийский, 70 | Процветают морские беспозвоночные. Широкое распространение трилобитов, водорослей. | ||
| Протерозойская (ранней жизни), свыше 2000 2700 | Органические остатки редки и малочисленны, но относятся ко всем типам беспозвоночных. Появление первичных хордовых - подтипа бесчерепных. | ||
| Архейская (самая древняя в истории Земли), около 1000. Возможно >3500 | Не заметны | ||
21. Предмет генетики, методы генетики.
Генетика—- наука о наследственности и изменчивости живых организмов и методах управления ими. В ее основу легли закономерности наследственности, установленные выдающимся чешским ученым Грегором Менделем (1822—1884) при скрещивании различных сортов гороха.
Наследственность — это неотъемлемое свойство всех живых существ сохранять и передавать в ряду поколений характерные для вида или популяции особенности строения, функционирования и развития. Наследственность обеспечивает постоянство и многообразие форм жизни и лежит в основе передачи наследственных задатков, ответственных за формирование признаков и свойств организма. Благодаря наследственности некоторые виды (например, кистеперая рыба латимерия, жившая в девонском периоде) оставались почти неизменными на протяжении сотен миллионов лет, воспроизводя за это время огромное количество поколений.
В то же время в природе существуют различия между особями как разных видов, так и одного и того же вида, сорта, породы и т. д. Это свидетельствует о том, что наследственность неразрывно связана с изменчивостью.
Изменчивость — способность организмов в процессе онтогенеза приобретать новые признаки и терять старые. Изменчивость выражается в том, что в любом поколении отдельные особи чем-то отличаются и друг от друга, и от своих родителей. Причиной этого является то, что признаки и свойства любого организма есть результат взаимодействия двух факторов: наследственной информации, полученной от родителей, и конкретных условий внешней среды, в которых шло индивидуальное развитие каждой особи. Поскольку условия среды никогда не бывают одинаковыми даже для особей одного вида или сорта (породы), становится понятным, почему организмы, имеющие одинаковые генотипы, часто заметно отличаются друг от друга по фенотипу, т. е. по внешним признакам.
Таким образом, наследственность, будучи консервативной, обеспечивает сохранение признаков и свойств организмов на протяжении многих поколений, а изменчивость обусловливает формирование новых признаков в результате изменения генетической информации или условий внешней среды.
Задачи генетики вытекают из установленных общих закономерностей наследственности и изменчивости. К этим задачам относятся исследования: 1) механизмов хранения и передачи генетической информации от родительских форм к дочерним; 2) механизма реализации этой информации в виде признаков и свойств организмов в процессе их индивидуального развития под контролем генов и влиянием условий внешней среды; 3) типов, причин и механизмов изменчивости всех живых существ; 4) взаимосвязи процессов наследственности, изменчивости и отбора как движущих факторов эволюции органического мира.
Генетика является также основой для решения ряда важнейших практических задач. К ним относятся: 1) выбор наиболее эффективных типов гибридизации и способов отбора; 2) управление развитием наследственных признаков с целью получения наиболее значимых для человека результатов; 3) искусственное получение наследственно измененных форм живых организмов; 4) разработка мероприятий по защите живой природы от вредных мутагенных воздействий различных факторов внешней среды и методов борьбы с наследственными болезнями человека, вредителями сельскохозяйственных растений и животных; 5) разработка методов генетической инженерии с целью получения высокоэффективных продуцентов биологически активных соединений, а также для создания принципиально новых технологий в селекции микроорганизмов, растений и животных.
При изучении наследственности и изменчивости на разных уровнях организации живой материи (молекулярный, клеточный,
организменный, популяционный) в генетике используют разнообразные методы современной биологии: гибридологический, цитогенетический, биохимический, генеалогический, близнецовый, мутационный и др. Однако среди множества методов изучения закономерностей наследственности центральное место принадлежит гибридологическому методу. Суть его заключается в гибридизации (скрещивании) организмов, отличающихся друг от друга по одному или нескольким признакам, с последующим анализом потомства. Этот метод позволяет анализировать закономерности наследования и изменчивости отдельных признаков и свойств организма при половом размножении, а также изменчивость генов и их комбинирование.
22. Закономерности наследования при моногибридном скрещивании, установленные Г. Менделем и их цитологическое обоснование. Промежуточное наследование.