
В СЛУЧАЕ СПИРАЛЬНОЙ СИММЕТРИИ, показанной на схеме строения вируса табачной мозаики, капсомеры, или субъединицы вируса, формируют спираль вокруг полой трубчатой сердцевины.








У отдельных бактериофагов (вирусов бактерий; фагов) смешанный тип симметрии. У т.н. "хвостатых" фагов головка имеет вид сферического капсида; от нее отходит длинный трубчатый отросток - "хвост".
КОМБИНИРОВАННАЯ, или смешанная, симметрия у вирусов может быть представлена разными вариантами. Частица бактериофага, показанная на схеме, имеет "головку" правильной геометрической формы и "хвост" со спиральной симметрией.
Встречаются вирусы с еще более сложным строением. Вирионы поксвирусов (вирусы группы оспы) не имеют правильного, типичного капсида: между сердцевиной и наружной оболочкой у них располагаются трубчатые и мембранные структуры.
РЕПЛИКАЦИЯ ВИРУСОВ
Генетическую информацию, закодированную в отдельном гене, в общем можно рассматривать как инструкцию по производству определенного белка в клетке. Такая инструкция воспринимается клеткой только в том случае, если она послана в виде мРНК. Поэтому клетки, у которых генетический материал представлен ДНК, должны "переписать" (транскрибировать) эту информацию в комплементарную копию мРНК
(см. также НУКЛЕИНОВЫЕ КИСЛОТЫ).
ДНК-содержащие вирусы по способу репликации отличаются от РНК-содержащих вирусов. ДНК обычно существует в виде двухцепочечных структур: две полинуклеотидные цепочки соединены водородными связями и закручены таким образом, что образуется двойная спираль. РНК, напротив, обычно существует в виде одноцепочечных структур. Однако геном отдельных вирусов представляет собой одноцепочечную ДНК или двухцепочечную РНК. Нити (цепочки) вирусной нуклеиновой кислоты, двойные или одинарные, могут иметь линейную форму или замыкаться в кольцо. Первый этап репликации вирусов связан с проникновением вирусной нуклеиновой кислоты в клетку организма-хозяина. Этому процессу могут способствовать специальные ферменты, входящие в состав капсида или внешней оболочки вириона, причем оболочка остается снаружи клетки или вирион теряет ее сразу после проникновения внутрь клетки. Вирус находит подходящую для его размножения клетку, контактируя отдельными участками своего капсида (или внешней оболочки) со специфическими рецепторами на поверхности клетки по типу "ключ - замок". Если специфические ("узнающие") рецепторы на поверхности клетки отсутствуют, то клетка не чувствительна к вирусной инфекции: вирус в нее не проникает. Для того чтобы реализовать свою генетическую информацию, проникшая в клетку вирусная ДНК транскрибируется специальными ферментами в мРНК. Образовавшаяся мРНК перемещается к клеточным "фабрикам" синтеза белка - рибосомам, где она заменяет клеточные "послания" собственными "инструкциями" и транслируется (прочитывается), в результате чего синтезируются вирусные белки. Сама же вирусная ДНК многократно удваивается (дуплицируется) при участии другого набора ферментов, как вирусных, так и принадлежащих клетке. Синтезированный белок, который используется для строительства капсида, и размноженная во многих копиях вирусная ДНК объединяются и формируют новые, "дочерние" вирионы. Сформированное вирусное потомство покидает использованную клетку и заражает новые: цикл репродукции вируса повторяется. Некоторые вирусы во время отпочковывания от поверхности клетки захватывают часть клеточной мембраны, в которую "заблаговременно" встроились вирусные белки, и таким образом приобретают оболочку. Что касается клетки-хозяина, то она в итоге оказывается поврежденной или даже полностью разрушенной. У некоторых ДНК-содержащих вирусов сам цикл репродукции в клетке не связан с немедленной репликацией вирусной ДНК; вместо этого вирусная ДНК встраивается (интегрируется) в ДНК клетки-хозяина. На этой стадии вирус как единое структурное образование исчезает: его геном становится частью генетического аппарата клетки и даже реплицируется в составе клеточной ДНК во время деления клетки. Однако впоследствии, иногда через много лет, вирус может появиться вновь - запускается механизм синтеза вирусных белков, которые, объединяясь с вирусной ДНК, формируют новые вирионы. У некоторых РНК-содержащих вирусов геном (РНК) может непосредственно выполнять роль мРНК. Однако эта особенность характерна только для вирусов с "+" нитью РНК (т.е. с РНК, имеющей положительную полярность). У вирусов с "-" нитью РНК последняя должна сначала "переписаться" в "+" нить; только после этого начинается синтез вирусных белков и происходит репликация вируса. Так называемые ретровирусы содержат в качестве генома РНК и имеют необычный способ транскрипции генетического материала: вместо транскрипции ДНК в РНК, как это происходит в клетке и характерно для ДНК-содержащих вирусов, их РНК транскрибируется в ДНК. Двухцепочечная ДНК вируса затем встраивается в хромосомную ДНК клетки. На матрице такой вирусной ДНК синтезируется новая вирусная РНК, которая, как и другие, определяет синтез вирусных белков.
См. также РЕТРОВИРУСЫ.
КЛАССИФИКАЦИЯ ВИРУСОВ
Если вирусы действительно являются мобильными генетическими элементами, получившими "автономию" (независимость) от генетического аппарата их хозяев (разных типов клеток), то разные группы вирусов (с разным геномом, строением и репликацией) должны были возникнуть независимо друг от друга. Поэтому построить для всех вирусов единую родословную, связывающую их на основе эволюционных взаимоотношений, невозможно. Принципы "естественной" классификации, используемые в систематике животных, не подходят для вирусов. Тем не менее система классификации вирусов необходима в практической работе, и попытки ее создания предпринимались неоднократно. Наиболее продуктивным оказался подход, основанный на структурно-функциональной характеристике вирусов: чтобы отличить разные группы вирусов друг от друга, описывают тип их нуклеиновой кислоты (ДНК или РНК, каждая из которых может быть одноцепочечной или двухцепочечной), ее размеры (число нуклеотидов в цепочке нуклеиновой кислоты), число молекул нуклеиновой кислоты в одном вирионе, геометрию вириона и особенности строения капсида и наружной оболочки вириона, тип хозяина (растения, бактерии, насекомые, млекопитающие и т.д.), особенности вызываемой вирусами патологии (симптомы и характер заболевания), антигенные свойства вирусных белков и особенности реакции иммунной системы организма на внедрение вируса. В систему классификации вирусов не вполне укладывается группа микроскопических возбудителей болезней, называемая вироидами (т.е. вирусоподобными частицами). Вироиды вызывают многие распространенные среди растений болезни. Это мельчайшие инфекционные агенты, лишенные даже простейшего белкового чехла (имеющегося у всех вирусов); они состоят только из замкнутой в кольцо одноцепочечной РНК.
5.Белки, их строение и функции.
Белки и их строение.
Среди органических компонентов клетки самыми важными являются белки. Они очень разнообразны и по строению, и по функциям. Содержание белков в различных клетках может колебаться от 50 до 80%.
Белки представляют собой высокомолекулярные (молекулярная масса до 1,5 млн углеродных единиц) органические соединения. Кроме С, О, Н, N, в состав белков могут входить S, Р, Fe. Белки построены из мономеров, которыми являются аминокислоты. Поскольку в состав молекул белков может входить большое число аминокислот, то их молекулярная масса бывает очень большой.
В клетках разных живых организмов встречается свыше 170 различных аминокислот, но бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Из них может быть образовано 2 432 902 008 176 640 000 комбинаций, т. е. различных белков, которые будут обладать совершенно одинаковым составом, но различным строением. Но и это огромное число не предел — белок может состоять и из большего числа аминокислотных остатков, и, кроме того, каждая аминокислота может встречаться в белке несколько раз.
Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (—NH2) с основными свойствами, другая — карбоксильной группой (—СООН) с кислотными свойствами. Часть молекулы, называемая радикалом (R), у разных аминокислот имеет различное строение (рис. 12).
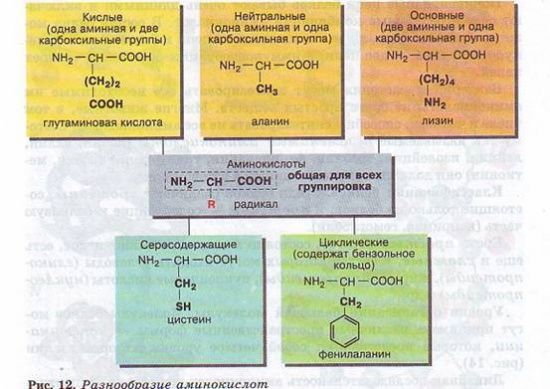
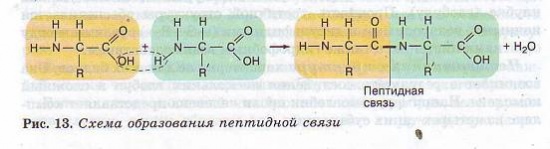
Наличие в одной молекуле аминокислоты и основной, и кислотной групп обусловливает их амфотерность и высокую реактивность. Через эти группы происходят соединения аминокислот при образовании белка. В ходе реакции полимеризации выделяется молекула воды, а освободившиеся электроны образуют ковалентную связь, которая получила название пептидной — образуется пептид (греч. peptos — сваренный). К свободным карбоксильной и аминогруппе могут присоединяться другие аминокислоты, удлиняя «цепь», называющуюся полипептидной. На одном конце такой цепи всегда будет группа МН2 (этот конец называется N-концом), а на другом конце — группа СООН (этот конец получил название С-конца) (рис. 13).
Полипептидные цепи белков бывают очень длинными и включают самые различные комбинации аминокислот. В состав белка может входить не одна, а две полипептидные цепи и более. Так, в молекуле инсулина — две цепи, а иммуноглобулины состоят из четырех цепей.
Бактерии и растения могут синтезировать все необходимые им аминокислоты из более простых веществ. Многие животные, в том числе и человек, способны синтезировать не все аминокислоты, поэтому так называемые незаменимые аминокислоты (лизин, валин, лейцин, изолейцин, треонин, фенилаланин, триптофан, тирозин, метионин) они должны получать с пищей в готовом виде.
Классификация белков.
Среди белков различают протеины, состоящие только из белков, и протеиды — содержащие небелковую часть (например, гемоглобин).
Кроме простых белков, состоящих только из аминокислот, есть еще и сложные, в состав которых могут входить углеводы (гликопротеиды), жиры (липопротеиды), нуклеиновые кислоты (нуклеопротеиды) и др.