
I комплекс – НАДН-дегідрогеназа (НАДН-убіхінон оксидоредуктаза) має дві функції: окислює НАДН.Н+ - переносить від нього 2 електрони на убіхінон та транспортує 4Н+ з матриксу у міжмембранний простір.








II комплекс – сукцинатдегідрогеназа (сукцинат- убіхінон оксидоредуктаза) забезпечує додатковий шлях для входу електронів у дихальний ланцюг за рахунок окиснення сукцинату.
III комплекс – цитохром с редуктаза (убіхінон-цитохром с оксидоредуктаза) містить цитохроми b та c 1, транспортує 2 електрони з убіхінону на цитохром с та забезпечує викачування 4 протонів з матриксу в міжмембранний простір.
IV комплекс – цитохром с оксидаза містить цитохроми а та а3, транспортує 2 електрони на кисень та одночасно переносить 2 протони в міжмембранний простір.
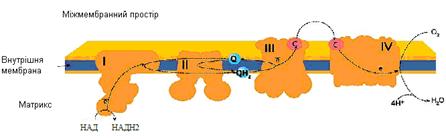
Рисунок 18 - Схема розташування комплексів дихального ланцюга
Робота дихального ланцюга приводить до синтезу молекул АТФ – це окисне фосфорилювання, механізми якого будуть розглянуті нижче. Між тим необхідно звернути увагу на те, що для синтезу 1 молекули АТФ потрібно приблизно 32 кДж/моль енергії. Така енергія виділяється в разі якщо різниця окисно-відновного потенціалу між переносниками в дихальному ланцюзі становить не менше 260 мВ.
Було встановлено, що в електрон-транспортному ланцюзі існують три ділянки з таким перепадом окисно-відновного потенціалу і вони відповідають комплексам I, II та IV. Ці ділянки були названі пунктами спряження з окисним фосфорилюванням, тобто при транспорті електронів через ці ділянки виділяється достатньо енергії для синтезу молекули АТФ (рис. 17).
Електрони, що транспортуються з НАДН.Н+, проходять три пункти спряження, тобто виділяється достатня кількість енергії для синтезу 3 молекул АТФ. Перенесення електронів з ФАДН2 відбувається лише через два пункти спряження (рис. 16), тому можливий синтез лише 2 молекул АТФ. Саме тому завжди реалізація енергії НАДН.Н+ супроводжується синтезом 3 АТФ, для ФАДН2 – 2 АТФ.
4.3. Інгібітори дихального ланцюга
Робота дихального ланцюга може бути інгібована під дією різноманітних клітинних отрут, які взаємодіють з білками-переносниками та порушують транспорт електронів:
- Інгібітори I комплексу — барбітурати (амітил, секонал), ротенон, пієрицидин А.
- Інгібітор II комплексу — малонат.
- Інгібітор III комплексу — антиміцин А.
- Інгібітори IV комплексу — Н2S, цианіди, СО.
Наявність у клітині вказаних інгібіторів призводить до розвитку гіпоенергетичного стану, що, в свою чергу, в деяких випадках може призвести до загибелі клітини.
4.4. Хеміосмотична теорія Мітчелла (теорія окисного фосфорилювання)
Пояснення механізмів спряження роботи дихального ланцюга та синтезу АТФ було запропоноване англійським біохіміком Пітером Мітчеллом у 1961 році, яке потім знайшло експериментальне підтвердження у роботах багатьох дослідни-ків. За винайдення механізмів окисного фосфорилювання П. Мітчелл у 1978 році отримав Нобелівську премії в галузі хімії («За внесок у розуміння процесу перенесення біологічної енергії, який зроблений завдяки створенню хеміосмотичної теорії»).
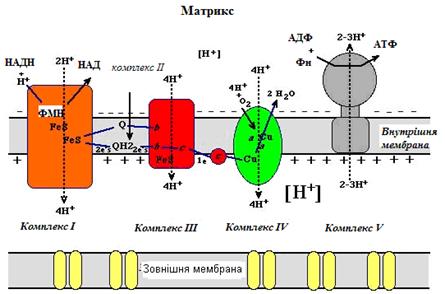
Рисунок 19 - Спряження роботи дихального ланцюга із синтезом АТФ (за теорією П.Мітчелла)
Основними постулатами теорії Мітчелла є таке (рис. 19):
1) внутрішня мембрана мітохондрій непроникна для іонів та малих молекул (за винятком молекул води);
2) дихальний ланцюг працює як «помпа», що викачує протони з матриксу в міжмембранний простір – рух 2 електронів від субстрату на кисень призводить до перенесення 8-10 Н+ (протони транспортуються через I, III та IV комплекси) через мембрану;
3) робота дихального ланцюга створює електрохімічний градієнт протонів (∆μН+), оскільки вони вільно через внутрішню мітохондріальну мембрану повернутися в матрикс не можуть і будуть накопичуватись у міжмемб-ранному просторі; ∆μН+ - це проміжна форма зберігання енергії окиснення субстратів;
4) енергію протонного градієнта використовує Н+-АТФ- синтаза (V комплекс) для синтезу АТФ, коли через одну з її субодиниць протони повертаються в матрикс;
5) існують сполуки - роз’єднувачі окисного фосфорилю-вання, які порушують електрохімічний градієнт протонів і знижують ефективність роботи Н+-АТФ-синтази.