
Синтез АТФ відбувається в ході реакцій двох основних процесів – окисного фосфорилювання та субстратного фосфорилювання.








Окисне фосфорилювання – процес синтезу АТФ при окисненні субстратів з участю дихального ланцюга мітохондрій (механізм процесу буде розглянутий нижче). Реакції цього процесу відбуваються виключно в аеробних умовах.
Субстратне фосфорилювання - процес синтезу АТФ, який відбувається як результат розщеплення субстратів без участі дихального ланцюга мітохондрій. У цьому разі перетво-рення субстрату в продукт супроводжується фосфорилюванням АДФ з утворенням АТФ. Цей процес можливий як в аеробних, так і анаеробних умовах.
На рис. 2 наведена формула АТФ, де вказані два макроергічних зв’язки в молекулі (~). Гідроліз кожного з них супроводжується виділенням енергії ∆Go≈ - 30 кДж/моль.
Виходячи з того, що молекула АТФ містить два високоенергетичні зв’язки, виділення енергії може відбуватися у вигляді двох реакцій:
АТФ → АДФ + Фн (∆Go= - 30 кДж/моль);
АТФ → АМФ + ФФн (∆Go= - 30 кДж/моль).
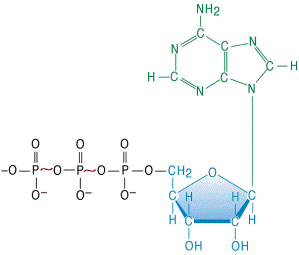
Рисунок 3 - Хімічна формула молекули АТФ
Тобто загальна кількість енергії, що утворюється при гідролізі АТФ, не може перевищувати - 30 кДж/моль.
Перша реакція найбільш поширена. Енергія зв’язків молекули АДФ, яка утворюється, не може бути використана, оскільки другий високоенергетичний зв'язок після відщеплення першої фосфатної групи буде перетворений у звичайний ковалентний зв'язок. Відомо, що подальший гідроліз АДФ та АМФ супроводжуються виділенням лише теплової енергії.
На рис. 4 схематично показані шляхи використання енергії АТФ в живих організмах.
2.2. Тканинне дихання та біологічне окиснення
Найбільш потужними процесами продукування енергії в клітинах є аеробні. Використання клітинами кисню необхідне для окиснення субстратів, кінцевими продуктами якого є Н2О, СО2, АТФ. Крім того, виділяється аміак, який у подальшому підлягає знешкодженню. Таким чином, клітини, тканини «дихають» і виробляють енергію, яка необхідна для життєдіяльності.
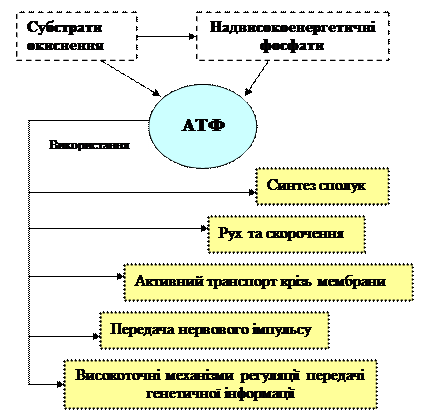
Рисунок 4 - Шляхи використання енергії АТФ

Тобто при тканинному диханні відбувається використання кисню для біологічного окиснення субстратів.
Між поняттями «тканинне дихання» та «біологічне окиснення» не слід ставити знак рівності, оскільки окиснення біосубстратів може відбуватися як в аеробних, так і у анаеробних умовах.

Основними субстратами біологічного окиснення є вуглеводи та ліпіди, саме їх катаболізм дає найбільшу кількість хімічної енергії, що акумулюється у високоенергетичних зв’язках макроергів. Білки також можуть бути використані з цією метою, але в організмі це не дуже «популярний» субстрат окиснення.
У тканинному диханні виділяють три стадії:
1) окиснення субстратів до ацетил-КоА;
2) розпад ацетильного залишку в ЦЛК;
3) транспорт електронів по дихальному ланцюгу міто-хондрій та сполучення цього процесу із синтезом АТФ.
Окиснення субстратів відбувається при:
1) відщепленні атома водню;
2) втраті електронів;
3) заміщення атомів на більш електронегативні атоми.
Окисно-відновні реакції, що відбуваються в процесі біологічного окиснення, каталізують ферменти класу оксидоредуктаз.
Виділяють три класи реакцій біологічного окиснення:
1. Реакції дегідрування.
У результаті цих реакцій відбувається перенесення водню з субстрату (S) на акцептор (А):
SH2 + A → S + AH2.
Ферменти, що каталізують рекції цього класу, мають назву дегідрогенази. Коферментами дегідрогеназ можуть бути НАД, НАДФ, ФАД, ФМН.
Залежно від типу акцептора водню реакції дегідрування поділяють на два підкласи:
- реакції, які каталізують анаеробні дегідрогенази. Акцептором (А) є сполука, яка відмінна від кисню:
SH2 + R → S + RH2;
- реакції, які каталізують аеробні дегідрогенази (оксидази).
Акцептором (А) в цих реакціях є кисень (О2):
SH2 + О → S + H2О2.
2. Реакції з перенесенням електрона/електронів:
Sē + А → S + Аē.
Такі реакції каталізують цитохроми (наприклад, цитохроми дихального ланцюга мітохондрій).
3. Оксигеназні реакції.
Реакції цього класу каталізують ферменти оксигенази, які приєднують до субстрату окиснення один або два атоми кисню. Тому залежно від кількості атомів кисню, що приєднуються, ферменти поділяють на:
- монооксигенази (відповідно реакції – монооксигеназні):
SH + 1/2О2 → S-ОН
До монооксигеназ належить цитохром Р450, що каталізує гідроксилювання ендогенних та екзогенних субстратів;
- диокисгенази (відповідно реакції – диоксигеназні):
S + О2 → SО2.
До диоксигеназних реакцій належать реакції перекисного окиснення ненасичених жирних кислот, що входять до складу ліпідів біомембран (реакції ПОЛ – перекисного окиснення ліпідів).
2.2.1. Ферменти біологічного окиснення
Ферменти біологічного окиснення залежно від будови поділяють на три класи:
1. Піридинзалежні дегідрогенази – дегідрогенази, які містять нікотинамідні коферменти (НАД+ або НАДФ+) (нікотинамід, що входить до складу цих коферментів, належить до похідного піридину).
Схематично реакції, що каталізують ці ферменти, мають такий вигляд:
SH2 + НАД+→S + НАДН + Н+;
та
SH2 + НАДФ+→S + НАДФН + Н+.