
Различают два вида тетануса: зубчатый и гладкий. Гладкий тетанус воз-никает при такой стимуляции мышцы, когда каждый последующий импульс








31
ГЛАВА 2. Физиология возбудимых тканей
возбуждения поступает к ней в фазу укорочения, а зубчатый - в фазу рассла-бления (рис. 9).
Амплитуда тетанического сокращения превышает амплитуду одиночного сокращения. Исходя из этого Гельмгольц объяснил процесс тетанического со-кращения простой суперпозицией - суммацией амплитуды одного мышечного сокращения с амплитудой другого. Однако в дальнейшем было показано, что при тетанусе имеет место не только сложение двух механических эффектов, т. к.
в определенных условиях амплитуда тетануса может быть больше или меньше суммы двух складывающихся сокращений. Н. Е. Введенский объяснил это яв-ление изменениями возбудимости мышцы, введя понятия оптимума и пессиму-ма частоты раздражения. Оптимальной называется такая частота раздражения, при которой каждое последующее раздражение наносится на мышцу в фазу повышенной возбудимости. При повышении возбудимости мышца способна отвечать на раздражение большей амплитудой и длительностью сокращения. Тетанус при этом будет максимальным (оптимальным), что проявляется в боль-шем количестве выполняемой работы. Пессимальной называется такая часто-та раздражения, при которой каждое последующее раздражение наносится на мышцу в фазу пониженной возбудимости. Тетанус при этом будет по амплиту-де минимальным или меньше ожидаемого (пессимальным), что проявляется в уменьшении объема выполняемой работы.
Режимы .мышечных сокращений. Различают изотонический, изометри-ческий и смешанный режимы сокращения мышц.
При изотоническом сокращении мышцы происходит изменение ее длины,
а напряжение остается постоянным. Такое сокращение возникает при отсут-ствии нагрузки на мышцу (например, при раздражении изолированной мышцы лягушки, закрепленной одним концом на штативе). В естественных услови-ях близкими к изотоническому типу сокращений являются сокращения мышц языка.
При изометрическом сокращении длина мышечных волокон остается по-стоянной, меняется лишь степень их напряжения. Такое сокращение мышцы можно получить созданием сопротивления, сила которого превышает силу мышечного сокращения (например, при поднятии непосильного груза). Этот режим наблюдается при сохранении заданной позы и при выполнении стати-ческой работы.
В целом организме сокращения мышц, как правило, имеют смешанный характер, т. е. происходит одновременное изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим, если преоб-ладает напряжение мышцы, или ауксометрическим, если преобладает укоро-чение.
32
Физиология мышц.
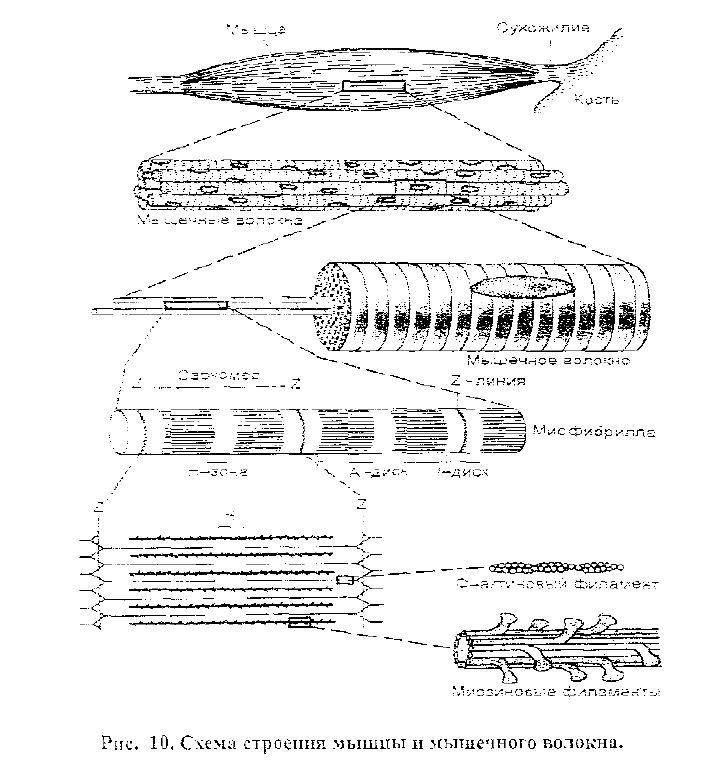
Механизм мышечного сокращения.
Скелетные мышцы состоят из мышечных волокон, которые, в свою оче-редь. состоят из множества тонких нитей - дшофибрилл, расположенных продольно. Каждая миофибрилла состоит из протофибрилл - нитей сократи-тельных белков: актина или миозина (рис. 10). Поперечные перегородки, на-зываемые Z-плистинами (или Z - мембранами), разделяют миофибриллы и, соответственно, мышечное волокно на участки - саркомеры. В саркомере на-блюдают поперечные правильно чередующиеся светлые и темные полосы. Эта поперечная исчерченность миофибрилл обусловлена определенным располо-жением нитей актина и миозина. В центральной части саркомера расположены толстые нити миозина, а в обоих концах саркомера находятся тонкие нити акти-на, прикрепленные к Z-пластинам. Центральная часть саркомера, содержащая только миозиновые протофибриллы. в световом поляризационном микроскопе выглядит как светлая полоска (Н-зона) в темном диске, который называется
33
ГЛАВА 2. Физиология возбудимых тканей
анизотропным или А-диском. Эта часть саркомера содержит нити и актина, и миозина, что и вызывает двойное преломление луча света. По обе стороны от
А-диска находятся участки, которые содержат только тонкие нити актина. Т.
к. они вызывают только одно лучепреломление света, то кажутся светлыми, и называются изотропными или J-дисками. По их середине проходит темная линия — Z-мембрана. Именно благодаря такому периодическому чередованию светлых (J) и темных (А) дисков сердечная и скелетные мышцы выглядят по-перечно исчерченными (поперечно-полосатыми).
В состоянии покоя концы толстых и тонких нитей лишь незначительно перекрываются на уровне А-диска. В соответствии с теорией скользящих ни-тей, при сокращении тонкие актиновые нити скользят вдоль толстых миози-новых нитей к середине саркомера, уменьшая его длину или (и) увеличивая напряжение. При этом актиновые и миозиновые протофибриллы своей длины не меняют.
Механизм скольжения нитей. Миозиновые нити имеют на своих концах выступы (поперечные мостики), утолщенные на конце и направленные в сто-рону активных нитей (рис. 10). Это утолщение называется головкой попереч-ного мостика. Актиновая нить состоит из двух закрученных одна вокруг дру-гой цепочек (подобно скрученным ниткам бус) глобулярных молекул актина. На нитях актина расположены молекулы тропонина, связанные с нитями белка тропомиозина. Тропомиозиновая нить в состоянии покоя мышцы располагает-ся в желобках между двумя молекулами актина так, что предотвращает прикре-пление головок поперечных мостиков миозина к активным центрам актиновых протофибрилл.
Во многих местах поверхностная мембрана мышечного волокна углубля-ется в виде микротрубочек внутрь волокна, перпендикулярно его продольной оси, образуя поперечные Т-трубочки. Параллельно миофибриллам и перпен-дикулярно Т-трубочкам между миофибриллами расположена система продоль-ных цистерн (саркоплазматический ретикулум). Концевые (терминальные) расширения этих цистерн подходят очень близко к поперечным трубочкам, об-разуя совместно с ними, так называемые, триады (Т-системы). В цистернах сосредоточено основное количество внутриклеточного кальция.
В состоянии покоя миозиновый мостик заряжен энергией (фосфорилиро-ван), но он не может соединиться с нитью актина, так как между ними нахо-дится система из нитей тропомиозина и глобул тропонина. При возбуждении ПД распространяется по мембранам Т-трубочек внутрь мышечного волокна и вызывает высвобождение ионов кальция из цистерн саркоплазматического ретикулума в протофибриллярное пространство. Кальций взаимодействует с тропонином, изменяя его пространственную конфигурацию, что приводит к смещению нити тропомиозина и открытию активных участков молекул актина. Происходит соединение миозиновой головки с актином и диссоциация про-
34
дуктов гидролиза АТФ, изменение пространственного положения («сгибание») головки и в результате - перемещение нити актина на один шаг (на один «гре-бок») к середине саркомера. Затем АТФ снова связывается с головкой миозина, что вызывает отсоединение мостика от актина. Ритмические прикрепления и отсоединения «гребущих» головок миозина создают силу, которая перемещает актиновую нить и вместе с ней Z-мембрану к середине саркомера, что приво-дит к укорочению мышечного волокна или (и) увеличению его напряжения.
При отсутствии повторного импульса возбуждения концентрация ионов кальция в протофибриллярном пространстве уменьшается, т. к. они закачи-ваются кальциевым насосом в систему цистерн саркоплазматического рети-кулума с использованием энергии АТФ. Отсоединение кальция от тропонина приводит к восстановлению первоначального пространственного положения тропомиозина, он снова опускается в желобки, блокируя активные центры ак-тина. Одновременно происходит фосфорилирование миозина. Таким образом, молекулы АТФ заряжают энергией сократительные элементы для следующего цикла этого процесса, а также способствуют разобщению актиновых и мио-зиновых нитей и обеспечивают работу кальциевого насоса, принимая участие как в сокращении, так и в расслаблении мышцы. Удлинение (расслабление) мышцы после ее сокращения является процессом пассивным, поскольку акти- . новые протофибри.ллы легко скользят в обратном направлении под влиянием сил упругости сократившихся мышечных волокон и мышцы, а также растяги-вающих их сил сокращающихся мышц антагонистов.
Гладкие мышцы.
Гладкие мышцы, формирующие мышечные слои стенок желудка, кишеч-ника, мочеточников, бронхов, кровеносных сосудов и других полых внутрен-них органов, построены из веретенообразных одноядерных мышечных клеток (волокон). Они не имеют поперечной исчерченности, поскольку в них миофи-бриллы расположены не строго параллельно друг другу, а хаотично. Клетки в гладких мышцах функционально связаны между собой специальными струк-турами наружных мембран - щелевидными контактами {нексусами), имеющи-ми низкое электрическое сопротивление. За счет этих контактов потенциалы действия и локальные (медленные) потенциалы распространяются с одного мышечного волокна на другое. Поэтому, несмотря на то, что двигательные нервные окончания заканчиваются на небольшом числе мышечных волокон, в реакцию вовлекается вся мышца. Следовательно, гладкие мышцы представля-ют собой функциональный синцитий.
Важным свойством гладких мышц является их пластичность, т. е. спо-собность сохранять приданную им при растяжении или деформации форму без изменения напряжения. Скелетная мышца в норме почти не обладает пла-
35
ГЛАВА 2. Физиология возбудимых тканей
стичностью. Эти различия хорошо видны при растяжении гладкой и скелетной мышцы. При удалении растягивающего груза скелетная мышца быстро укора-чивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования органов. Так, на-пример, пластичность мышц мочевого пузыря по мере его наполнения мочой предотвращает значительное повышение давления в нем, что в свою очередь не препятствует оттоку мочи из лоханок почек и. тем самым, не нарушает процесс мочеобразования. Благодаря пластичности гладкая мышца может быть полно-стью расслаблена как в укороченном, так и в растянутом состоянии.
Гладкие мышцы обладают способностью к автоматии, обусловленной наличием специализированных «пейсмекерных» клеток. Они идентичны по структуре другим гладкомышечным клеткам, но отличаются по электрофизио-логическим свойствам. В пейсмекерных клетках возникает спонтанная мед-ленная деполяризация мембраны (препотенциал). При достижении критиче-ского уровня происходит дальнейшая деполяризация мембраны и развитие ПД. Восходящая часть спайка возникает, главным образом, за счет входа в клетку ионов кальция, при этом мембранный потенциал достигает значения +20 мВ. Затем деполяризация сменяется реполяризацией. и МПП восстанавливается. ПД длится несколько секунд. За реполяризацией следует новый препотенциал, который вызывает следующий потенциал действия и т. д. Частота спонтанных ПД определяет величину миогенного тонуса гладкой мышцы.
Особенностью гладких мышц является их способность осуществлять от-носительно медленные ритмические и длительные тонические сокращения. Медленные ритмические сокращения гладких мышц желудка, кишечника, мо-четочников и других полых органов обеспечивают перемещение их содержи-мого из одной области органа в другую. Длительные тонические сокращения гладких мышц, особенно сфинктеров полых органов, препятствуют свободно-му выходу из этих органов их содержимого или его свободному перемещению из одной области органа в другую. Гладкие мышцы стенок кровеносных со-судов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосу-дов влияет на величину их просвета и, следовательно, на уровень артериально-го давления крови и кровоснабжения органов.
Выраженность тонуса зависит от исходной длины гладкомышечных во-локон. Быстрое растяжение гладких мышц вызывает нарастающую деполяри-зацию мышечных клеток и их последующее сокращение. Сокращение, вызван-ное растяжением, играет важную роль в авторегуляции тонуса кровеносных сосудов, перемещении содержимого кишки, а также обеспечивает непроиз-вольное (автоматическое) опорожнение переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
36
Физиология мышц.
Активность пейсмекерных гладкомышечных клеток лежит в основе спон-танных тетанообразных сокращений гладких мышц. Т.к. в гладких мышцах одиночное сокращение продолжается несколько секунд, их тетанус возникает даже при низкой частоте их стимуляции. В отличие от скелетной мышцы глад-кие мышцы кишки, мочеточника, желудка и матки развивают спонтанные тета-нообразные сокращения. В этом случае сокращения возникают не в результате передачи импульсов возбуждения с нерва на мышцу, а вследствие активности мышечных клеток, обладающих автоматией.
Автономная нервная система и ее медиаторы оказывают на спонтанную активность пейсмекеров гладких мышц модулирующие влияния. Так, напри-мер, при нанесении ацетилхолина на препарат мышцы толстой кишки пейс-мекерные клетки деполяризуются до околопорогового (около критического) уровня, и частота ПД возрастает. Инициируемые ими сокращения сливаются, образуя почти гладкий тетанус. Чем выше частота ПД, тем сильнее суммиро-ванное сокращение. Норадреналин же гиперполяризует мембрану тех же кле-ток. в результате чего уменьшается частота генерации ПД, и следовательно, уменьшается величина тонуса мышцы.
Механизм сокращения гладких мышц. Возбуждение гладкомышечных клеток вызывает увеличение входа ионов кальция в клетку и высвобождение ионов кальция из ее саркоплазматического ретикулума. В результате повыше-ния концентрации ионов кальция в саркоплазме активируются сократительные структуры, но механизм активации их в гладком волокне отличается от меха-низма активации в поперечно-полосатом. В гладкомышечной клетке кальций взаимодействует с белком кальмодулином. Он активирует киназы легких цепей миозина, которые за счет АТФ фосфорилируют головки миозиновых мостиков. Они соединяются с активными центрами актиновой протофибриллы и совер-шают «гребок». Так же. как сердечная и скелетные мышцы, гладкие мышцы расслабляются пассивно. Однако расслабление гладких мышц происходит бо-лее медленно, т. к. в них кальциевый насос откачивает ионы кальция из цито-плазмы в саркоплазматический ретикулум и межклеточное пространство менее производительно.
37
Глава 3. Физиология центральной нервной системы
ГЛАВА 3. ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Центральная нервная система ~ это часть нервной системы позвоночных животных, представленная скоплением нервных клеток, образующих спинной и головной мозг.