
Механизм проведения возбуждения по нервным волокнам.








Механизм проведения возбуждения у различных нервных волокон прин-ципиально одинаков. Он осуществляется на основе ионных механизмов гене-рации потенциала действия. Вместе с тем, имеются и некоторые различия в механизмах проведения возбуждения по миелиновым и безмиелиновым волок-нам.
При распространении возбуждения по безмиелиновому нервному волокну (рис. 7а) местные электрические токи, которые возникают между возбужден-ным участком волокна (его поверхность заряжена отрицательно) и близлежа-щим невозбужденным (его поверхность заряжена положительно), вызывают деполяризацию мембраны до ее критического уровня с последующей генера-цией ПД на невозбужденном участке мембраны. В следующий момент локаль-ные токи возникают уже между только что возбужденной частью мембраны
и следующим ближайшим невозбужденным участком. Процесс генерации ПД
в этой части невозбужденной мембраны повторяется. Таким образом, проис-ходит распространение возбуждения вдоль нервного волокна без уменьшения
22
Физиология нервов и нервных волокон.
амплитуды ПД. Поскольку при этом в процесс возбуждения последовательно вовлекаются участки всей без исключения мембраны волокна, то такой меха-низм проведения возбуждения называется последовательным или непрерыв-ным.
Наличие у миелиновых волокон миелиновой оболочки, обладающей высо-ким электрическим сопротивлением, а также участков волокна, лишенных та-кой оболочки (перехваты Ранвъе) создает условия для качественно иного типа проведения возбуждения. Здесь местные электрические токи возникают не
.между соседними участками мембраны, а между соседними перехватами Ран-вье, разделенными участком волокна, покрытым миелиновой оболочкой (рис. 76). Следовательно, возбуждение как бы «перепрыгивает» через эти участки от одного перехвата к другому. Такой механизм распространения возбуждения на-зывается салыпаторным (скачкообразным) или прерывистым. Скорость тако-го способа проведения возбуждения значительно выше, и он более экономичен по сравнению с непрерывным, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
«Перепрыгивание» потенциала действия через межперехватный участок, покрытый миелиновой облолочкой, возможно потому, что ПД создает местные токи, величина которых в 5-6 раз превышает необходимую для возбуждения соседнего перехвата. ПД может «перепрыгивать» не только через один, но и че-рез два - четыре межперехватных промежутка. Это явление может наблюдать-ся, например, при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества (например, новокаина).
Лабильность и парабиоз нервных волокон. Нервные волокна, как и дру-гие возбудимые структуры, обладают лабильностью, которая определяется длительностью спайка потенциала действия, т. е. длительностью фазы абсо-лютной рефрактерности. Так как она у спайкового потенциала нервного во-локна самая короткая, то и лабильность последнего самая высокая. Нервное волокно способно воспроизводить до 1000 импульсов в секунду.
Н. Е. Введенский обнаружил, что при альтерации участка нерва путем воздействия на него повреждающего агента (например, анестетика или охлаж-дения) происходит снижение его лабильности. Оно наступает в результате за-медления процессов восстановления исходного функционального состояния альтерированного участка волокна после каждого прохождения через него им-пульса возбуждения. В основе этого явления на первых этапах лежит процесс инактивации натриевой проницаемости мембраны, к которому затем присоеди-няется и нарушение калиевой проницаемости. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом, а фактор, его вызвав-ший - парабиотическим агентом. В развитии парабиоза выделяют три фазы, последовательно сменяющих друг друга: уравнительную, парадоксальную, тормозную.
23
ГЛАВА 2. Физиология возбудимых тканей
В уравнительную фазу происходит уравнивание величины ответной реак-ции возбудимой структуры на действие как слабых, так и сильных раздражите-лей. Например, при низкой частоте раздражения нервного волокна (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как его возбудимость после прохождения каждого импульса успевает вос-становиться. При высокой частоте раздражений (100 Гц) импульсы могут по-ступать к альтерированному участку в тот момент, когда он еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенци-алом действия. Поэтому часть импульсов по нерву к мышце не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100), то величина ответной реакции мышцы становится такой же, как и на ред-кие раздражители (25 Гц).
В парадоксальную фазу происходит дальнейшее снижение лабильности. При этом на сильные раздражители ответная реакция значительно меньше, чем на слабые (наблюдается парадокс), т. к. сильные раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности.
В тормозную фазу лабильность снижается до такой степени, что и слабые,
и сильные раздражители не вызывают ответной реакции. При этом мембрана нервного волокна находится в состоянии длительной деполяризации в резуль-тате потери способности к реполяризации, т. е. к восстановлению исходного функционального состояния.
В организме, как правило, сила раздражителя кодируется частотой возбуж-дений. Поэтому описанные закономерности развития парабиоза для сильных и слабых раздражители справедливы для частых и редких воздействий.
Парабиоз - явление обратимое. Если парабиотический агент действует не-долго, то после прекращения его действия нерв выходит из состояния парабио-за через те же фазы, но в обратной последовательности.
Физиология синапсов.
Синапс — это специализированная структура, которая обеспечивает пере-дачу сигналов от одной возбудимой структуры к другой. Термин «синапс» был введен Ч. Шеррингтоном 1897 году и означает «сведение», «соединение», «за-стежка».
Классификация синапсов. Синапсы можно классифицировать по разным критериям:
1) их местоположению и принадлежности соответствующим структурам:
• периферические (нервно-мышечные, нейро-секреторные и др.);
• центральные (аксо-соматические, аксо-дендритные, аксо-аксональные); 2) функции - возбуждающие и тормозные;
24
Физиология синапсов.
3) механизму передачи сигналов - химические, электрические, смешан-
ные.
4) медиатору, с помощью которого осуществляется передача сигнала в хи-мическом синапсе - холинергические, адренергические, серотонинергические, глицинергические и т. д.
Передача сигнала осуществляется: в химических синапсах - посредством выделения и действия химического вещества (медиатора), в электрических - за счет возникновения и действия электрического тока, в смешанных - с ис-пользованием обоих этих механизмов. У человека и высших животных в нерв-ной системе преобладают синапсы с химическим способом передачи возбуж-дения.
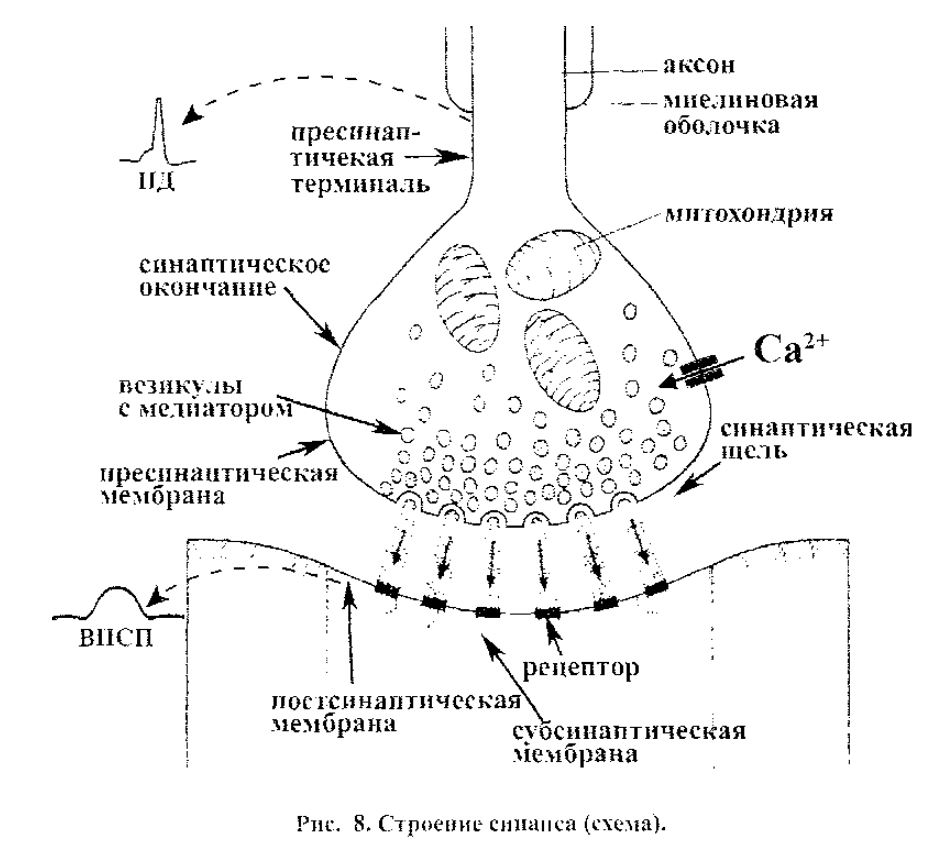
Строение химического синапса. Синапс состоит из следующих основ-ных элементов (рис.8):
• синаптической бляшки с синаптическими везикулами,
• пресинаптической мембраны,
• постсинаптической мембраны.
• синаптической щели.
25
ГЛАВА 2. Физиология возбудимых тканей
Синоптическая бляшка представляет собой расширенное окончание нерв-ного волокна (чаще - аксона), внутри нее находятся синаптические везикулы
- пузырьки, содержащие медиатор. Пресинаптическая мембрана - это часть мембраны нервного окончания в области его синаптического контакта с другой клеткой. В ней локализованы потенциалзависимые кальциевые каналы, необ-
ходимые-для активации выделения медиатора. Постсинаптическая мембрана
- часть мембраны клетки, воспринимающей возбуждение, в области синапти-
ческого контакта. Участок постсинаптической мембраны, расположенный на-против пресинаптической (непосредственно под ней), называется субсинапти-ческой мембраной. Особенностью субсинаптической мембраны является нали-чие в ней специальных рецепторов, чувствительных к определенному медиа-тору и связанных с хемозависимыми ионными каналами. Существуют два типа рецепторов: ионотропные, белковая структура которых является одновременно ионным каналом, и метаботропные, взаимодействие которых с медиатором приводит к изменению обменных процессов в клетке.
В постсинаптической мембране, за пределами субсинаптической, локали-зуются только потенциалозависимые каналы. Синаптическая гцелъ - это меж-клеточное пространство шириной 40-50 нм между пре- и постсинаптическими мембранами, заполненное межклеточными жидкостью и веществом.