
3. Следовая деполяризация - отражает процесс медленной реполяризации мембраны.








15
ГЛАВА 2. Физиология возбудимых тканей
4. Следовая гиперполяризация - отражает увеличение поляризации мем-браны относительно исходного уровня. Гиперполяризация мембраны в этот период обусловлена, во-первых, продолжающимся выходом ионов калия; во-вторых, возможно, открытием каналов для хлора и поступлением этих ионов в цитоплазму клетки; в-третьих, усиленной работой натрий-калиевого насоса.
Изменение возбудимости при возбуждении. Развитие любого вида воз-буждения (местного или распространяющегося) сопровождается фазным из-менением уровня возбудимости, обычно оцениваемого по порогу раздражения ткани. Характер этих изменений и их соотношение с фазами одиночного цикла возбуждения (потенциала действия), иллюстрирует рис. 4Б.
Состоянию исходной поляризации мембраны в покое соответствует ис-ходное состояние ее возбудимости или нормальный уровень возбудимости, принимаемый за 100%.
В период развития предспайкового потенциала возбудимость повышается. Эта фаза изменения возбудимости получила название фазы первичной экзаль-тации. Механизм повышения возбудимости в этот период связан с приближе-нием процесса деполяризации к его критическому уровню. В этих условиях сила раздражителя, необходимая для доведения процесса до КУД, будет мень-шей, т. к. уменьшается прочность закрытия активационных ворот натриевых каналов.
В период развития спайкового потенциала натриевые каналы открыты полностью, в результате чего мембрана утрачивает способность отвечать на действие раздражителя любой, даже очень большой силы. Эта фаза называется фазой абсолютной рефрактерности (абсолютной невозбудимости).
В период реполяризации исходное состояние натриевых каналов мембра-ны постепенно восстанавливается, что сопровождается сменой абсолютной рефрактерности мембраны на относительную рефрактерность, т. е. ответная реакция может быть, но для ее возникновения необходимо использовать сверх-пороговые раздражители.
Периоду следовой деполяризации соответствует повышенный уровень возбудимости — фаза вторичной экзальтации (супернормальной возбудимо-сти). Так как мембранный потенциал в эту фазу ближе к КУД по сравнению с состоянием покоя (исходной поляризацией), то порог раздражения снижен, и новое возбуждение в эту фазу может возникнуть при действии раздражителей подпороговой силы. В период развития следовой гиперполяризации возбуди-мость ткани понижена - фаза вторичной рефрактерности (субнормальной воз-будимости). В эту фазу мембранный потенциал увеличивается (состояние ги-перполяризации мембраны), удаляясь от критического уровня деполяризации. Следовательно, порог раздражения повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины.
16
Законы раздражения возбудимых тканей.
Законы раздражения возбудимых тканей.
Законы раздражения отражают определенную зависимость между дей-ствием раздражителя и ответной реакцией возбудимой ткани.
Закон «силы»: величина ответной реакции ткани на раздражение пря-мо пропорциональна силе раздражителя (до определенного уровня). В соот-ветствии с этим законом функционируют сложные целостные структуры, на-пример, скелетная мышца. С увеличением силы раздражителя амплитуда ее сокращений постепенно увеличивается от минимальных (пороговых) величин до субмаксимальных и максимальных значений. Это обусловлено тем, что ске-летная мышца состоит из множества мышечных волокон, имеющих разную возбудимость. Поэтому на пороговый раздражитель отвечают только те мы-шечные волокна, которые имеют самую высокую возбудимость - амплитуда мышечного сокращения при этом минимальна. С увеличением силы раздра-жителя по мере достижения порога раздражения в реакцию вовлекается все большее количество мышечных волокон, и амплитуда сокращения мышцы уве-личивается. Когда в реакцию вовлечены все мышечные волокна, составляю-щие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения, и наблюдающееся сокращение называют максимальным.
Закон «все или ничего»: величина ответной реакции ткани на раздра-жение не зависит от силы раздражителя - при подпороговом раздражении возбуждение (ПД) не возникает («ничего»), а при пороговом и сверхпороговых раздражениях всегда возникает реакция максимальной амплитуды («все»). По закону «все или ничего» на раздражитель реагируют одиночные нервное и мы-шечное волокна, а также сердечная мышца.
Закон «все или ничего» не абсолютен. Во-первых, на раздражители подпо-роговой силы в ткани происходят невидимые изменения, получившие название местного возбуждения (локального ответа), которые проявляются, прежде все-го, в изменении мембранного потенциала покоя. Во-вторых, сердечная мышца, растянутая кровью заполнившей полости сердца, реагирует также по закону «все или ничего», но амплитуда ее сокращения будет большей по сравнению с сокращением сердечной мышцы, не растянутой кровью.
Закон физиологического электротона Э. Пфлюгера: действие постоян-ного тока на ткань сопровождается изменениями ее возбудимости. Началь-ное прохождении постоянного тока через нерв или мышцу в участке ткани, расположенном под катодом повышает возбудимость (катэлектротон), а под анодом - понижает (анэлектротон). Изменения возбудимости обуславливают-ся частичной деполяризацией клеточной мембраны под катодом и гиперполя-ризацией под анодом. Эти изменения возбудимости получили название элек-тротона (электротоническое изменение возбудимости).
17
ГЛАВА 2. Физиология возбудимых тканей
При дальнейшем действии постоянного тока первоначальное повышение возбудимости под катодом сменяется ее понижением (катодическая депрес-сия). Первоначальное же снижение возбудимости под анодом сменяется ее по-вышением (анодная экзальтация). Развитие католической депрессии связанно
с инактивацией натриевой проницаемости мембраны, а анодной экзальтации
- с уменьшением калиевой проницаемости и ослаблением исходной натриевой инактивации.
Закон полярного действия постоянного тока Э. Пфлюгера: при замы-кании цепи постоянного тока возбуждение возникает под катодом, а при раз-мыкании — под анодом. Это связано с тем, что прохождение постоянного элек-трического тока через возбудимую ткань вызывает изменение мембранного потенциала покоя ее клеток. Так, под катодом положительный заряд наружной поверхности клеточной мембраны уменьшается - начальная деполяризация мембраны, которая затем под влиянием проходящего электрического тока бы-стро достигает критического уровня и вызывает распространяющееся возбуж-дение. Под анодом положительный заряд наружной поверхности мембраны возрастает - гиперполяризация мембраны, но при этом критический уровень деполяризации смещается к уровню потенциала покоя. При размыкании цепи тока гиперполяризация мембраны исчезает, ее потенциал возвращается к ис-ходной величине и достигает смещенного критического уровня деполяризации
- возникает возбуждение.
Закон раздражения Э. Дюбуа-Рейлюна: раздражающее действие тока зависит не только от его абсолютной величины, но и от скорости его изме-нения во времени. Пороговая сила тока увеличивается при снижении крутизны его нарастания (рис. 5).
Существует некоторое минимальное значение скорости нарастания силы раздражителя, при которой ответная реакция вообще не возникает, даже при неограниченном увеличении силы раздражителя. Это явление получило на-звание аккомодации. В его основе лежит изменение состояния потенциалза-висимых натриевых и калиевых каналов клеточной мембраны. При быстром увеличении силы стимула повышение натриевой проницаемости мембраны обеспечивает достижение критического уровня деполяризации прежде, чем наступит инактивация натриевых каналов. При медленном нарастании силы раздражителя натриевые каналы переходят в состояние инактивации по мере нарастания деполяризации, что снижает возбудимость мембраны вплоть до полной рефрактерности.
Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника и желудка.
Закон силы - длительности Г. Вейсса -II. Лапика: раздражающее дей-ствие постоянного тока зависит не только от его абсолютной величины, но
18
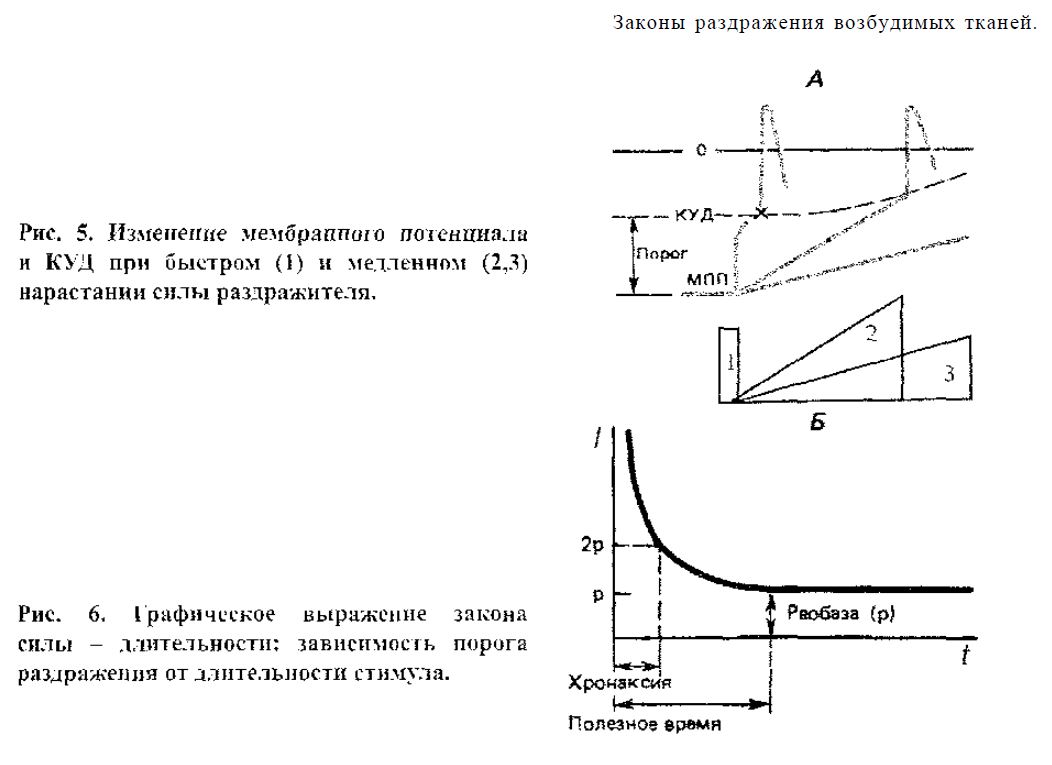
и от времени, в течение которого он действует на ткань. Чем больше сила раздражителя, тем меньше времени он должен действовать для возникновения возбуждения.
Этот закон вскрывает гиперболическую зависимость пороговой силы раз-дражителя от длительности его действия (рис. 6). Ток, сила которого ниже не-которой минимальной величины, не вызовет возбуждения, как бы длительно он ни действовал. С другой стороны, чем короче импульс тока, тем меньшую раздражающую способность он имеет. Раздражающий ток, длительность кото-рого меньше 0,01 мс, не вызывает ответной реакции при любой силе стимула.
В основе такой закономерности является неспособность раздражителя осуществить сдвиг мембранного потенциала до КУД из-за кратковременности действия или из-за недостаточности силы раздражителя.
Минимальная величина тока, способная вызвать возбуждение при нео-граниченно длительном его действии, называется реобазой. Время, в течение которого действует и вызывает возбуждение ток, равный реобазе, называется полезным временем. В связи с тем, что определение этого времени затруднено, было введено понятие хронаксии. Хронаксия - минимальное время, в течение которого должен действовать ток, равный двум реобазам, чтобы вызвать от-ветную реакцию. Определение хронаксии методом хронаксиметрии находит применение в клинике. Электрический ток, приложенный к мышце, проходит
19
ГЛАВА 2. Физиология возбудимых тканей
как через мышечные, так и через нервные волокна, находящиеся в этой мыш-це. Хронаксия нервных волокон значительно меньше хронаксии мышечных волокон. При повреждении нерва или гибели соответствующих мотонейронов спинного мозга (например, при полиомиелите) происходит разрушение нерв-ных волокон. В этом случае определяется хронаксия мышечных волокон, кото-рая имеет большую величину.