
Воротные каналы имеют «ворота», которые открываются и закрываются








в ответ на изменения МПП (такие каналы называются потенциалзависимыми), или в ответ на действие химических веществ (хемозависимые или лигандзави-симые каналы), или при деформации мембраны (механочувствительные кана-лы). В покое большинство воротных каналов закрыто.
По способности пропускать различные ионы (ионной избирательности) каналы подразделяют на моноселективные, обладающие способностью про-водить преимущественно один вид ионов; полиселективные, пропускающие в равной степени несколько видов ионов; неселективные, пропускающие боль-шое число молекул разных типов.
Принцип устройства и работы воротных каналов можно рассмотреть на примере потенциалзависимых. Селективные потенциалзависимые ионные ка-налы подразделяются на натриевые, калиевые, кальциевые и хлорные. Канал состоит из собственно канала (транспортной части) и воротного механизма («ворот»), который управляется электрическим полем мембраны. В натриевых каналах предполагают наличие двух типов «ворот»: быстрых или активаиион-ных ( т ) и медленных или инактивагшонных (h) (рис.3). «Ворота» могут быть открыты или закрыты. Например, в состоянии покоя клетки в натриевом канале «ворота» m закрыты, а «ворота» h открыты. При уменьшении заряда мембраны (деполяризации) «ворота» m открываются, и канал переходит в проводящее со-стояние, т. е. через него начинают проходить ионы натрия под действием сил концентрационного и электрохимического градиентов. Затем, при достижении
12
Биоэлектрические явления в живых тканях. Природа возбуждения.
процессом деполяризации определенного момента закрываются инактиваци-онные «ворота», и канал перестает пропускать ионы, т. е. он инактивируется. По мере восстановления МПП (реполяризации) инактивационные «ворота» медленно открываются, а активационные быстро закрываются, и канал воз-вращается в свое исходное функциональное состояние. В других каналах есть только активационные ворота.
В состоянии покоя, при наличии трансмембранного градиента концентра-ций ионов происходит, прежде всего, выход калия из клетки в межклеточное пространство по каналам утечки. Выход положительно заряженных ионов ка-лия приводит к формированию положительного заряда на наружной поверх-ности мембраны. При этом органические анионы (отрицательно заряженные крупномолекулярные соединения), для которых мембрана клетки непроницае-ма, придают внутренней поверхности мембраны отрицательный заряд. На сте-пень поляризации мембраны в состоянии покоя оказывает влияние перемеще-ние через нее и других ионов (натрия, хлора) по каналам различных типов, но оно в этих условиях невелико. Например, мембрана нервных волокон в состоя-нии покоя в 25 раз менее проницаема для ионов натрия, чем для ионов калия.
Перемещение ионов через мембрану по их концентрационным градиен-там в конечном счете должно было бы привести к выравниванию их концен-трации внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране функционируют ионные насосы. Так, важное значение для поддержания МПП имеет натрий-калиевый насос, обеспечивающий выведение из цитоплазмы клетки ионов натрия и введение
в цитоплазму ионов калия. Ионный насос перемещает ионы против их кон-центрационного градиента и. следовательно, работает с затратой энергии на преодоление силы градиента. При этом насос переносит через мембрану нео-динаковое количество зарядов (три иона натрия в обмен на два иона калия), что усиливает поляризацию мембраны.
Выходу избытка калия из клетки также препятствует электрическое поле, возникающее при накоплении положительного заряда снаружи клетки и отри-цательного - внутри.
Таким образом, возникновение и поддержание МПП покоя обусловлено избирательной проницаемостью мембраны клетки для ионов и работой натрий-калиевого насоса.
Мембранный потенциал покоя создает электрическое поле, которое под-держивает закрытое состояние активационных «ворот» и открытое состояние инактивационных «ворот» натриевых каналов, а также способствует сохране-нию определенной пространственной организации мембраны.
Регистрация электрических потенциалов в нервном, мышечном волокнах или в нервной клетке показала, что при действии раздражителя происходит изменение МПП. Под влиянием раздражителя подпороговой силы изменения
13
ГЛАВА 2. Физиология возбудимых тканей
МПГТ невелики и имеют местный (локальный) характер. Такое изменение МПП получило название локального потенциала, и оно не сопровождается целост-ной реакцией клетки. Но при действии раздражителя пороговой или сверхпо-роговой силы изменения мембранного потенциала максимальны и последова-тельно охватывают всю мембрану клетки. Это изменение получило название распространяющегося потенциала или потенциала действия (ПД), поскольку он вызывает характерную для данной клетки специализированную реакцию.
Механизм возникновения ПД заключается в следующем. Под влиянием раздражителя пороговой или сверхпороговой величины проницаемость мем-
14
Биоэлектрические явления в живых тканях. Природа возбуждения.
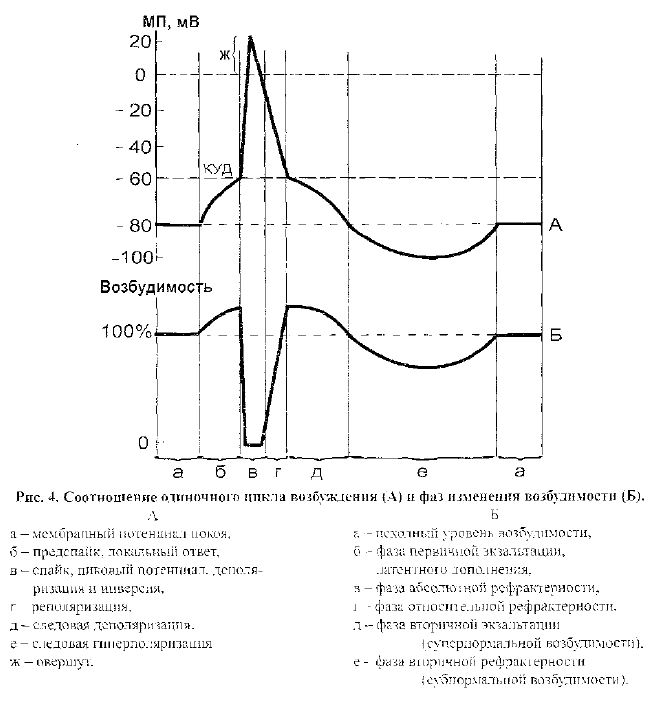
ораны клетки для ионов натрия постепенно возрастает. Вход ионов натрия в клетку приводит к уменьшению величины мембранного потенциала покоя - де-поляризации мембраны (рис. 4А). Вначале процесс деполяризации развивается медленно, и возникает предспайковый, медленный, локальный потенциал. При уменьшении мембранного потенциала до критического уровня деполяризации (КУД) открывается достаточное количество потенциалзависимых натриевых каналов, и вход ионов натрия в клетку начинает превышать выход ионов калия, что приводит к дальнейшей деполяризации и открытию остальных натриевых каналов. В результате проницаемость мембраны для ионов натрия быстро, ла-винообразно увеличивается (в 500 раз) и превышает проницаемость для ионов калия в 20 раз. Это приводит к развитию ПД и формированию спайкового или быстрого потенциала. При развитии восходящей части спайкового потенциала вследствие проникновения ионов натрия в цитоплазму и их взаимодействия с отрицательно заряженными анионами трансмембранная разность потенциалов сначала исчезает, а затем происходит перезарядка клеточной мембраны (по-ляризация с обратным знаком, или реверсия заряда), когда внутренняя поверх-ность мембраны заряжается положительно по отношению к наружной. Этот потенциал превышения (овершут) достигает величины +30 - +50 мВ. На этом уровне потенциала начинают закрываться инактивационные ворота быстрых натриевых каналов (происходит инактивация натриевой проницаемости), и от-крываются калиевые каналы, через которые ионы калия выходят из клетки. Это приводит к восстановлению исходного уровня мембранного потенциала покоя,
т. е. происходит реполяризация мембраны. Процесс реполяризации отражают нисходящая часть спайкового потенциала, следовая деполяризация и следовая гиперполяризация.
Потенциал действия может быть зарегистрирован двумя способами: вне-клеточным - с помощью электродов, приложенных к внешней поверхности клетки; внутриклеточным - с помощью электродов, один из которых введен внутрь клетки, а другой расположен на ее поверхности.
При монополярном отведении от наружной поверхности клетки потен-циал действия имеет сложную форму и состоит из нескольких компонентов
(рис.4А).
1. Предспайковый потенциал (локальный ответ) - он отражает процесс медленной деполяризации мембраны от уровня МПП до критического уровня деполяризации.
2. Спайковый потенциал (спсшк, распространяющийся потенциал или соб-ственно потенциал действия) - своей восходящей частью отражает процесс быстрой деполяризации мембраны и ее перезарядки, а нисходящей - процесс быстрой реполяризации мембраны.