
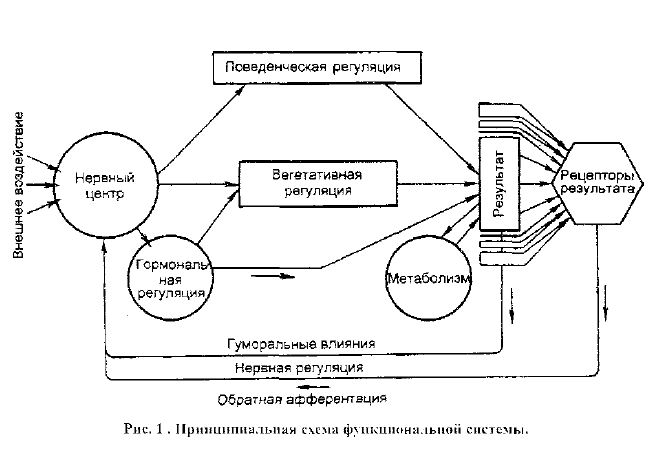
Аппараты контроля - рецепторы результата - образования (структуры), реагирующие на изменение состояния данного параметра, вызванное воздей-ствием внешних или внутренних факторов.








Аппараты управления (регуляции) - совокупность центральных структур, осуществляющих направленное влияние на деятельность аппаратов реакции (органов), от которых зависит восстановление исходного (нормального, кон-стантного) уровня отклонившегося параметра.
Аппараты реакции - органы и системы органов, изменение уровня функ-ционирования которых в соответствии с регулирующими влияниями аппаратов управления приводит к восстановлению исходной величины параметра.
Обратная афферентагшя - нервный и (или) гуморальный канал передачи информации в аппараты управления о достижении или не достижении полез-ного результата.
В основном все ФУС можно разделить на два типа: гомеостатические и поведенческие. Гомеостатические ФУС обеспечивают автоматическое под-держание параметров (констант) внутренней среды организма на относительно постоянном уровне. Если же их ресурсов недостаточно для восстановления ве-личин параметра, то формируется поведенческий компонент гомеостатической ФУС. Поведенческие ФУС обеспечивают формирование таких видов поведе-ния, которые связаны с удовлетворением доминирующей потребности.
Теория функциональных систем является важным инструментом в пони-мании закономерностей организации процессов саморегуляции, того или иного вида приспособительной деятельности организма и ее нарушений. При забо-
6
 левании человека анализ компонентов функциональной системы нарушенной деятельности помогает врачу наиболее эффективно осуществить поиск причин нарушения, его локализацию и степень выраженности, а затем наметить пути восстановления или компенсации нарушенной функции посредством методов врачебного воздействия на организм.
левании человека анализ компонентов функциональной системы нарушенной деятельности помогает врачу наиболее эффективно осуществить поиск причин нарушения, его локализацию и степень выраженности, а затем наметить пути восстановления или компенсации нарушенной функции посредством методов врачебного воздействия на организм.
Организм человека обладает выраженной способностью адаптироваться к постоянно меняющимся условиям внешней среды. В основе приспособитель-ных реакций организма лежит универсальное свойство живого объекта (клет-ки, ткани, органа) раздражимость — способность отвечать на действие раз-дражающих факторов изменением структурных и функциональных свойств, изменением обмена веществ. Раздражимостью обладают все ткани животных
и растительных организмов. В процессе эволюции происходила постепенная дифференциация тканей. При этом раздражимость некоторых из них достигла наивысшего развития и трансформировалась в новое свойство - возбудимость. Этим термином обозначают способность ткани отвечать на раздражение спец-ифическим изменением обмена веществ, специализированной реакцией - воз-буждением. Возбуждение - это специализированная ответная реакция возбуди-мого объекта на действие раздражителя, проявляющаяся в изменении обмен-ных процессов и генерации электрических потенциалов.
Возбудимостью обладают нервная, мышечная и железистая ткани. Их объ-единяют понятием «возбудимые ткани». Для них специализированными ответ-
7
ГЛАВА 1. Основные понятия физиолог ии
ными реакциями будут, соответственно, генерация и проведение возбуждения, сокращение, секреция. Возбудимость различных тканей неодинакова. Мерой возбудимости является порог раздражения - минимальная сила раздражителя, которая способна вызвать возбуждение. Менее сильные раздражители назы-ваются подпороговымн, а более сильные - сверхпороговыми. Раздражителем живого объекта может быть любое изменение внешней или внутренней среды организма, если оно достаточно велико, возникло достаточно быстро и продол-жается достаточно долго.
Классификация раздражителей. Все раздражители по их природе мож-но разделить на три группы:
• физические (механические, термические, электрические, звуковые, све-товые);
• химические (щелочи, кислоты, гормоны, медиаторы, продукты обмена веществ);
• физико-химические (изменение осмотического давления, рН среды, ион-ного состава).
По степени приспособленности реакции биологических объектов к дей-ствию раздражителя, все раздражители делятся на адекватные и неадекватные. Адекватными называются те из них, к действию которых в процессе эволюции биологический объект был приспособлен в наибольшей степени. Например, адекватным раздражителем для фоторецепторов является видимый свет, для барорецепторов - изменение давления, для скелетной мышцы - нервный им-пульс и т. д. Неадекватными называются такие раздражители, которые дей-ствуют на структуру, специально не приспособленную для реагирования на их действие. Например, адекватным раздражителем для скелетной мышцы явля-ется нервный импульс, но мышца может возбуждаться и при действии элек-трического тока, механического удара и др. Эти раздражители для скелетной мышцы являются неадекватными, и их пороговая сила в сотни или тысячи раз превышает пороговую силу адекватного раздражителя.
Лабильность (физиологическая подвижность) — способность воспро-изводить определенное количество циклов возбуждения в единицу времени в точном соответствии с ритмом действующих раздражителей. Мерой лабильно-сти является максимальное количество циклов возбуждения, которое способ-но воспроизвести возбудимое образование в единицу времени без трансфор-мации ритма раздражения. Свойство лабильности необходимо учитывать при действии на ткани ритмическими раздражителями.
8
Биоэлектрические явления в живых тканях. Природа возбуждения.
ГЛАВА 2. ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Биоэлектрические явления в живых тканях. Природа возбуждения.
Первые попытки последовательной разработки учения о «животном элек-тричестве» связаны с именем Л. Гальвани (1792 г.). Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенного на медном крючке, при их прикосновении к железным перилам балкона. На основании этих наблюдений Л. Гальвани пришел к выводу, что сокращение мышц лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается к ним по металлическим проводникам. Этот опыт в настоящее вре-мя известен как первый опыт Гальвани (опыт с металлами).
Физик А. Вольта, повторив опыт Гальвани, пришел к заключению, что опи-санные явления не связаны с «животным электричеством». Источником тока, по его мнению, являлся не спинной мозг, как полагал Л. Гальвани, а разность потенциалов, образующаяся в месте контакта разнородных металлов - меди и железа. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исклю-чив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал его при помощи стеклянного крючка на мышцы голени. При этом иногда возникало сокращение мышц. Этот опыт известен как второй опыт Гальвани или опыт без металлов.
Позже было выявлено, что сокращение мышц во втором опыте Гальвани возникало только тогда, когда нерв одновременно соприкасался с их повреж-денной и неповрежденной поверхностями. Э. Дюбуа-Реймоном было установ-лено, что поврежденный участок мышцы имеет отрицательный заряд, а непо-врежденный - положительный. При набрасывании нерва на поврежденный и неповрежденный участки мышцы возникает электрический ток, который раз-дражает нерв и вызывает сокращение мышцы. Этот ток был назван током по-коя или током повреждения.
Э. Дюбуа-Реймон. таким образом, впервые показал, что наружная поверх-ность мышцы заряжена положительно по отношению к ее внутреннему со-держимому. Следовательно, в состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая затем была названа мембранным потенциалом покоя (МПП). В современных условиях его измерение производится путем погружения микроэлектрода в ци-топлазму клетки и регистрации разности потенциалов между внутренней и на-ружной поверхностями мембраны. Величина МПП у разных клеток составляет от -60 до -90 мВ.
Для объяснения механизма возникновения и поддержания мембранного потенциала покоя было разработано несколько теорий. В 1949-52 гг. А. Ход-жкин, А. Хаксли, Б. Катц модифицировали существовавшие ранее гипотезы
9
ГЛАВА 2. Физиология возбудимых тканей
и экспериментально обосновали основные положения меморанно-ионнои теории. Согласно этой теории возникновение МПП обусловлено неодинако-вой концентрацией ионов натрия, калия, кальция, хлора внутри клетки и во внеклеточной среде, а также неодинаковой проницаемостью для этих ионов поверхностной мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 12 раз меньше ионов хлора, чем внеклеточная жидкость. Следовательно, в со-стоянии покоя существует асимметрия концентрации ионов внутри клетки и в окружающей ее среде.
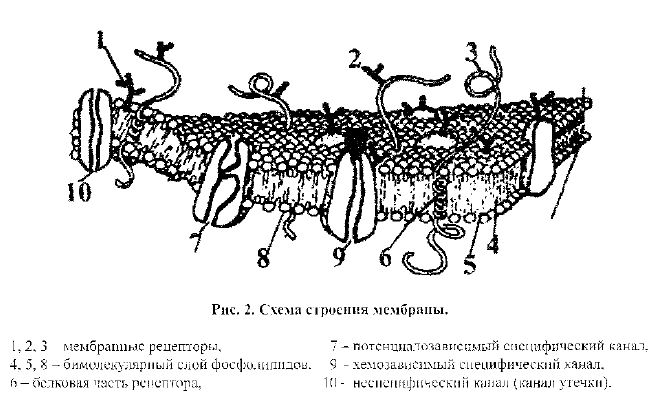
Клетку ограничивает тончайшая оболочка - поверхностная мембрана клетки, или плазматическая мембрана, или плазмалемма. В ее состав входят липиды (в основном фосфолипиды), белки (в основном гликопротеины) и углеводы (в основном мукополисахариды). Согласно общепринятой трехмер-ной жидкостно-мозаичной модели мембраны, она имеет следующую структуру (рис.2). Основу мембраны образует двойной слой липидов, в который погру-жены молекулы белков полностью (т. е. пронизывая его насквозь) или частич-но (интегральные белки). Другая группа мембранных белков, которые также могут быть частично погруженными в мембрану, но в основном связаны с ее внешней и внутренней поверхностями, называется поверхностными белками.
Липиды выполняют в основном функцию матрикса (основы) мембраны. Функции белков мембраны более многочисленны и разнообразны. Так, инте-гральные белки выполняют транспортную функцию, образуя ионные каналы
и переносчики веществ через мембрану; рецепторную, являясь рецепторами
10
Биоэлектрические явления в живых тканях. Природа возбуждения.
химических раздражителей (гормонов, медиаторов, антигенов); а также могут быть ферментами. Функции периферических белков еще более многочислен-ны: рецепторная. ферментативная, структурная, сократительная, адгезивная, медиаторная (использование в качестве посредников передачи сигнала между клетками).
Проницаемость наружной мембраны клетки для различных веществ определяется, в основном, наличием в ее составе транспортных белков, обра-зующих каналы для ионов или других веществ и различные переносчики или ионные насосы. Перенос веществ через мембрану может осуществляться с по-мощью пассивного или активного транспорта.
Активным называется транспорт вещества против концентрационного или электрохимического градиента с затратой клеткой энергии и с участием специ-альных переносчиков. Различают первично-активный и вторично-активный транспорт. Первично-активным транспортом называется процесс переноса вещества против концентрационного или электрохимического градиента с непосредственным использованием энергии клеточного метаболизма. Мем-бранные переносчики, обеспечивающие первично-активный транспорт, по сути представляют собой ферменты АТФ-азы, и называются насосами. При-мером такого вида транспорта является перенос ионов натрия и калия, который происходит при участии натрий-калиевого насоса с использованием энергии АТФ. Вторично-активным транспортом называется перенос вещества против градиента без непосредственной затраты энергии, за счет электрохимического градиента другого вещества (иона). С помощью такого механизма, например, происходит реабсорбция глюкозы в почках. При этом глюкоза из просвета ка-нальца входит в клетку стенки капилляра вместе с ионом натрия с помощью специального переносчика.
Пассивным называется транспорт вещества по концентрационному, осмо-тическому или электрохимическому градиентам без затраты энергии расщепле-ния АТФ и реализуется с участием мембранных каналов или по механизму об-легченной диффузии. Пассивный транспорт включает в себя диффузию, филь-трацию, осмос. Движущей силой диффузии частиц растворенного вещества является их концентрационный градиент. Разновидностью диффузии является осмос, при котором перемещение происходит в соответствии с концентраци-онным градиентом частиц растворителя. Под филыпрагщей понимают процесс переноса раствора через пористую мембрану под действием гидростатическо-го давления.
Облегченная диффузия, как и простая диффузия, осуществляется без за-траты энергии по градиенту концентрации. Однако облегченная диффузия представляет собой более быстрый процесс и осуществляется с участием пере-носчика.
11
ГЛАВА 2. Физиология возбудимых тканей
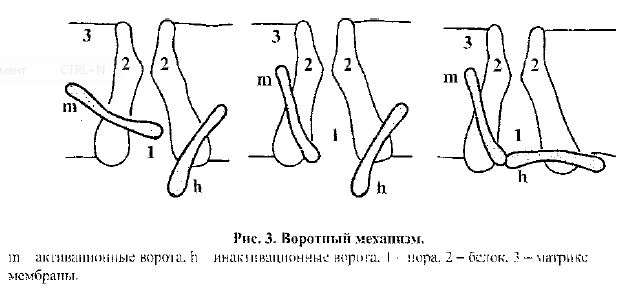
Ионные каналы мембраны делятся на каналы утечки и воротные (управля-емые). Каналы утечки не имеют ворот, открыты в покое, их состояние обычно не зависит от уровня мембранного потенциала, однако оно может изменяться при действии некоторых физических и химических факторов (например, ане-стетиков, температуры, рН и др.).