
5. - Метод пространственно-временных координат акустических сигналов.








6. - Фотографирование следов жизнедеятельности.
7. - Опрос.
В Воронежском заповеднике вся работа проводилась традиционными методами пространственной ориентации и привязки, основанными на высокоразвитой лесотехнической инфраструктуре заповедника, которая, к тому же, находилась в хорошей сохранности. Расстояние между квартальными столбами по широте равно 500 м, по долготе - 1000 м. Нумерация кварталов хорошо читается.
В «Калужских засеках» и «Орловском Рис. 6. След передней левой
|
|

конечности прибылого.
полесье» исследования проводились на фоне полноценного геоинформационного обеспечения. Это значит, что все значимые для нас маршруты и ориентиры находились в памяти приемника GPS и были в постоянном распоряжении исследователя. Помимо физического ознакомления с местностью, мы могли в любой момент проконсультироваться с электронными картами и при желании загрузить конкретные ориентиры на GPS.
IV.а.1. Методологический комплекс тропления
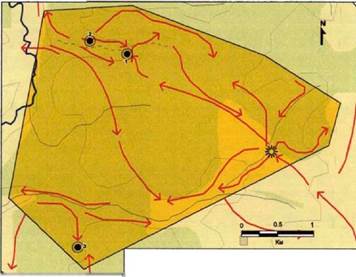
С самого начала работы метод тропления sensu lato был разделен на две взаимодополняющие части, обе основанные на анализе следов жизнедеятельности: тропление sensu stricto и "сканирование" пространства с помощью сети маршрутов. Эта сеть была разработана после ознакомления со структурой полигона, с его топографией, квартальной сетью и другими техническими аспектами, на основе которых и строилось освоение нами места проведения исследований. Такого рода освоение "места работы" представляется нам совершенно необходимым, так как физически и эмоционально вводит исследователя в среду.
Первый сезон для каждого полигона был посвящен прояснению общих вопросов, таких как количество волков, структура группирования внутри стаи (стай), часто используемые волками переходы, известные научным сотрудникам заповедников и национального парка. Для этого мы обосновывались в центральной части каждого полигона (Крутовской кордон - в Воронежском заповеднике; Ягодненский кордон - в «Калужских засеках»; Михайловский кордон - в «Орловском полесье»), с тем, чтобы иметь свободный доступ к областям, где к тому времени была отмечена наибольшая плотность встреч с волками или со следами их жизнедеятельности. Зимой мы приняли в качестве маршрутов все наши перемещения по вышеуказанным районам, как вдоль просек, косовых дорог, так и по целине, которые не следовали непосредственно за передвижением животных. Так, главное отличие проведения маршрутов от тропления состоит в обхвате большего пространства за единицу времени в первом случае, и в более высоком качестве информации, получаемой во втором случае. Маршруты охватывали как
|
Рис. 7. Сканирующие маршруты на территории Воронежского заповедника. |
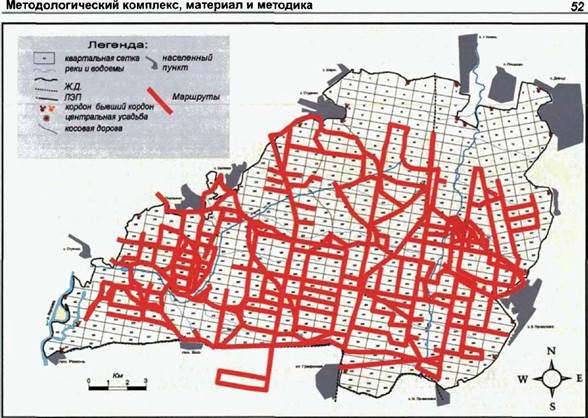
можно большую площадь для того, чтобы создать как бы «одномоментный снимок» картины распределения волков в пространстве. Совокупность сезонных маршрутов планировалась заранее таким образом, чтобы обследовать полигон максимально равномерно и во времени и в пространстве в течение каждого сезона (см. рис.7,8 и 9). В случае Воронежского заповедника каждый маршрут охватывал по меньше мере одну треть исследуемого участка; как в «Калужских засеках», так и в «Орловском полесье» - две трети. Любой выход в лес предполагал продвижение в направлении определенного, запланированного заранее района, до обнаружения следов жизнедеятельности волка.
Следующие за обнаружением следов действия определялись тем, какой вопрос ставился в начале тропления; в большинстве случаев мы придерживались заранее разработанного плана, будь то сам процесс тропления или процесс получения информации от него: в некоторых случаях больший интерес представляло ясное отражение передвижения животных, и тогда тропление проводилось словно в ускоренном темпе, что, в свою очередь, ограничивало возможности уточнения деталей поведения. В других случаях оказывалось более предпочтительным уделить повышенное внимание не столько всей протяженности
|
Рис. 8. Сканирующие маршруты на южном участке заповедника «Калужские засеки». |
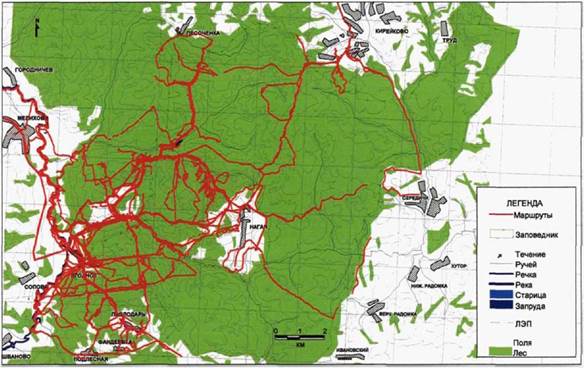
следа, сколько важным формам поведения, которые могли играть решающую роль. Мы редко вносили изменения в структуру наших планов, происходящие изменения были вызваны редкими событиями, такими, как обнаружение жертвы или присутствие животных в неизвестных районах и т.п.
|
Рис. 9. Сканирующие маршруты в национальном парке «Орловское полесье». |
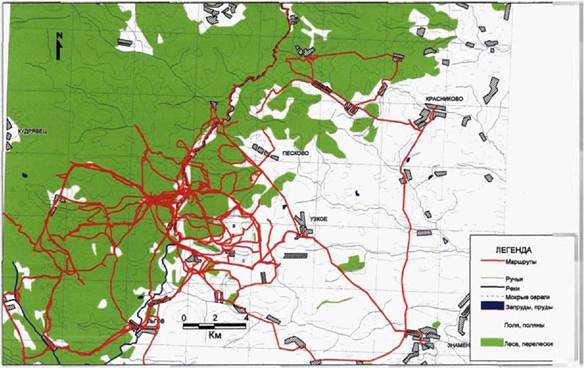
местоположения объектов велась с помощью шагомера, учитывая азимут и расстояние до ближайших квартальных столбов.
Во время тропления и продвижения по маршрутам тщательно собирались следующие данные:
/. Параметры отпечатков (следов): (см. ниже главу IV.a.2: Комплекс
индивидуальной идентификации особей: Подометрия) 2. Длина шага и аллюр. Эти параметры позволяли не только оценить ритм передвижения животного, но и уточнить, принадлежат ли обнаруженные на одной и той же дороге следы животным, передвигавшимся вместе или по отдельности.
1. Картографирование волчьих магистральных троп с помощью GPS или с занесением на карту их расположения, ориентации (в градусах), частоты использования (в случае, если это возможно).
2. Картографирование экскрементов с помощью GPS или с занесением на карту их расположения, степени сохранности и ее длительности, в случае неоднократного прохождения по одному и тому же маршруту. Кроме того, регистрировалось их содержание.
3. Картографирование мочевых меток с помощью GPS или с занесением на карту их расположения, а также типа меток: RLU - метка самца с поднятой лапой, FLU - метка самки с поднятой лапой, SQU - уринация на четырех лапах (сокращение по Питерсу и Мичу (1975)).
4. Картографирование поскребов с помощью GPS или с занесением на карту их
|
Рис. 11. Типы поскребов: а — со сходящимися сериями; б— с параллельными сериями; в — с расходящимися сериями. |

| б |
| в |
| а |
(МесИ, 1966). Для решения вопросов, касающихся социальной и пространственной организации, необходима идентификация исследуемых особей. Большинство попыток индивидуального опознавания особей лесных популяций, основанных только на визуальных признаках, не увенчалось успехом или, если они оказались результативными, требовало огромных усилий со стороны исследователя (Бадридзе, 1979). В последние сорок лет существовал только один способ идентификации и прослеживания волков, живущих не в открытых пространствах - радиослежение (МесЬ, 1983). В поисках других ответов на этот вопрос мы решили пересмотреть возможности классических методов и, в итоге, пришли к компромиссному варианту, основанному на взаимном дополнении классических и современных методов. Придерживаясь нашей доктрины методологических комплексов, мы включили в наш арсенал алгоритм идентификации, базирующийся на тщательном анализе следов жизнедеятельности волков, результаты которого указывают на высокую разрешающую способность данного методологического комплекса.
Подометрия
При проходе через высокую траву, мох и, особенно, влажную почву, песок, грязь и, наконец, снег, волки всегда оставляют следы. Сохранность этих следов зависит от метеоусловий. Размеры параметров следов конкретной конечности конкретной особи сохраняются в определенных пределах, обусловленные аллюром, весом животного, субстратом. Существование этих пределов описано в литературе (Матюшкин, 2000). Известно, что такие параметры, как длина и ширина следа, у собачьих могут позволить нам судить о возрасте и половой принадлежности животного (Формозов, 1954). Но можно ли применять промеры следов в качестве индивидуального идентификатора особей? Логика подсказывает, что разброс промеров по субстратам и при большом количестве особей позволит лишь обрисовать определенную кластерносгь, где каждый кластер будет отличаться от других только по принципу половых и возрастных категорий. Казалось бы, этим вопрос исчерпан. И действительно, многие авторы на этом и остановились (например, КагапЛ, 1987). Но что, если ограничить до определенного предела количество идентифицируемых особей и набор следообразующих субстратов? При
| Таблица 2. Статистическая обработка данных подометрии волков западной и восточной групп в Воронежском заповеднике (1997-99 гг.). Субстраты: снег, грязь и песок. Е - восточная группировка; \Л/- западная группировка; АО - передняя правая лапа; I - общая длина без когтей; А - общая ширина; Г - коэффициент Формозова (Ы/А&
|
| самцов самок |
| 700 300 |
| 1,14 ±0,01[1] 1,28 ±0,02* |
|
Рис. 12. След передней правой конечности волка. I. - длина; А - ширина. |
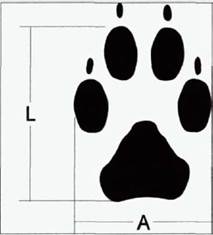
| таких условиях можно оценить количественно степень перекрывания вышеупомянутых пределов каждой из особей. Поставленный таким образом вопрос кажется нам вполне разрешимым. Для того, что дать на него четкий ответ, нужно лишь промерять достаточно большие выборки следов заранее индивидуально распознанных особей. Мы именно так и поступили в своей работе. Для индивидуального распознавания применяли метод кинологической идентификации, описанный ниже. |
Таблица 5. Статистическая обработка данных подометрии волков группы «Зуевка», на северной части национального парка «Орловское полесье» (2000-01 гг.). Субстраты: снег, грязь и песок.
| 62 |
| Методологический комплекс, материал и методика |
АО - передняя правая лапа; ¡. - общая длина без когтей; А - общая ширина; f - коэффициент Формозова (Ь^УА)
|
|
| (99-104) (88-90) (90-92) (95-96) (93-94) (76-80) (83-85) (87-88) (80-82) (84-86) (80-81) |
100,91 ± 0,20 89,50 ±0,12 90,67 ±0,11 95,38 ±0,08 93,33 ±0,13 77,76 ±0,32 84,00 ±0,23 87,75 ±0,10 81,00 ±0,33 84,67 ±0,25 80,50 ±0,17
AD, мм
А_
| Кличка Пол N |
| I 1Д2 1.27 1Д2 1,16 1,07 1,10 1.9 1,14 1,23 1.28 1.10 |
| Атар Келта Партизан Риз Дор Пир Ал Цид Лейка Алера |
| 55 50 60 40 15 21 20 20 10 15 10 |
| (85-95) (65-74) (78-85) (80-85) (80-92) (69-75) (72-81) (75-80) (64-68) (66-67) (72-75) |
| самец самка самец самец самец самец самец самец самка самка самец |
90,27 ±0,52 70,80 ±0,47 81,00 ±0,25 82,38 ± 0,32 88,00 ±1,51 70,95 ±0,51 77,50 ±0,77 77,25 ± 0,44 66,00 ±0,67 66,33 ±0,13 73,50 ±0,50
| 241 85 |
| 1,11 ±0,002* 1,26 ±0,006* |
| самец самка |
|
|
* - достоверность различий самцов и самок по «Ь, р«0,001 (критерий Манна-Уитни)
Таблица 6. Достоверность различий промеров длины отпечатков передних правых конечностей волков стаи «Зуевка», северная часть национального парка «Орловское полесье» (Критерий Манна-Уитни)

| Атар Келта Партизан Риз Дор Пир Ал Цид Лейка Алера |
| Щ р<0,001 NS |
пахучих образцов на примере других видов млекопитающих проводится уже в течение ряда лет (Соколов и др., 1990). Образцы мочи волков были использованы нами впервые (Крутова и др., 1999).
Образцы запаха волков собирали в полевых условиях по унифицированной методике (Сулимов, Поярков, 2001), без прямого контакта рук с меткой, для чего использовались чистые, т.е. не контаминированные предыдущими метками, предметы. Образцы меток заворачивали в хлопчатобумажную салфетку и в алюминиевую фольгу и хранили при минусовых температурах в индивидуальных герметических полиэтиленовых контейнерах с соответствующей этикеткой. В лабораторных условиях приготовление образцов запаха для предъявления собакам и их хранение велось согласно описанной ранее методике (Крутова, 1999). Идентификация образцов проводилась двумя способами - поиск по подобию и поиск по различию. В первом случае сравнение проводили стандартным методом выборки, при котором собаке предлагали запомнить пахучий образец, после чего проводили ее вдоль селективного ряда, составленного из образцов мочи волков группировки, к которой относился запоминаемый, так и других зверей, служивших контрольными образцами (волки из других стай и из зоопарка). Всего в селективном ряду имелось 10 проб. Собака последовательно их обнюхивала и, встретив образец, идентичный запомненному ранее, ложилась или садилась возле него (такое поведение вырабатывалось дрессировкой). Для того, чтобы оценить правильность работы собаки, одним из 10 образцов был «тестовый» - полученный от того же зверя, что и запоминаемый. Собака, ведущая адекватный поиск, садилась возле такого образца, что служило тестом ее нормального рабочего поведения. В результате анализа тестовый образец сопоставлялся с уже имеющимися и определенными. Таким образом, все запаховые образцы распределялись на группы, и каждая группа соответствовала индивиду, оставившему запаховые метки, с которых брались тестируемые образцы. В распознавании образцов по принципу поиска отличия сравнивали образцы в подборке из 10 объектов, большая часть которых (7-8) принадлежала особям одного пола, оставшиеся 2-3 - другого. В этом случае, как и в первом варианте, сигналом собаки об обнаружении ею искомых отличий служила посадка.
В экспериментах применяли трех подготовленных собак породы кеесхонд. Было проведено 474 опыта. Количество использованных образцов мочи и
Аппаратура и программное обеспечение
Для картографирования данных спутниковой навигационной системой использовались до четырех 12-канальных GPS еТгех и один 12-канальный GPS 12XL фирмы «Garmin».
Информация GPS в полевых условиях обрабатывалась с помощью компьютерной программы OziExplorer v. 3,90.3а. на портативном компьютере Toshiba satellite 4010CDS.
Для измерения следов использовался штангенциркуль и циркуль с линейкой. Расстояния во время тропления в Воронежском заповеднике измерялось при помощи шагомера Soligor РМ-520,
Ориентация (в градусах) измерялась при помощи геологического компаса фирмы «Tucuman».
Информация о троплении и маршрутах, а также визуальные наблюдения записывались на диктофон Sony TCM-359V,
Акустическая активность регистрировалась при помощи магнитофона Sony D-3 и узконаправленного микрофона Senhaizer.
Температура воздуха измерялась цифровым термометром фирмы «Sweden SRT AB», показывающим также минимальную и максимальную температуру данного интервала времени.
Атмосферное давление измерялось при помощи барометра фирмы «Сагас». Для визуальных наблюдений использовались бинокли Nikon 8x42 DCF HP и БПЦ5 8x30 и подзорная труба Halleyscope 8-32х 40.
Во время троплении, чтобы поддерживать связь при расходах, использовались выносные рации фирмы Midland модель Alan 95 plus.
При фотографировании мы пользовались объективами Nikkor AF24 мм f/2,8D, Nikkor 35-80 мм НА-5.6D, Sigma AF70-300 мм Аро macro и Tokina AF400 мм f/5.6 на корпусах фотоаппарат SLR Nikon Fl 00 и SLR Nikon F70. Использованная пленка: Kodachrome E-100S, Kodachrome E-100SW, Kodachrome Panther lOOx, 400x и pl600x, Agfachrome RSX 100, 200 и 50.
В качестве транспортного средства использовался внедорожник BA3-21213 и три лошади-полукровки.
контролирует количество вариации каждой составляющей данного расчета. Отсюда, чем меньше h, тем больше разрешающая способность метода и точность оценка данных.
Второй тип Кернела - адаптивный. В этом случае коэффициент сглаживания варьирует таким образом, что зоны с большей концентрацией точек имеют более высокие значения h, а в тех зонах, где концентрация точек меньше, h - меньше. Адаптивный кернел является усовершенствованной версией фиксированного. Вортон (1989) предлагает практический математический способ расчета идеального h в зависимости от размера выборки и от неравномерности и плотности распределении точек. Формула для определения оптимального значения h с фиксированными кернелами выглядит следующим образом:
где а ={1/2[о2х(1) + <^Х(2)]}1/2, и a2X(i),<^x(2) — предполагаемые отклонения данных х
лгт У*
На основе такого математического аппарата строился компьютерный модуль, используемый нами в данной работе.
Для анализа данных методами «Минимального полигона» и «Кернела» использовался компьютерный программный модуль Animal movements v.2.0 для пакета геоинформационной системы Arc View 3.2а компании «Esri».
Критерии выделения пространственных субъединиц
Выделение границ таких пространственных субъединиц семейного участка волков, как очаг и жизненное пространство, проводилось с помощью метода «минимального полигона» по следующим критериями:
Жизненное пространство - для определения границ этой пространственной субъединицы мы использовали точки регистраций охотничьего поведения родительской пары вместе с прибылыми.
Очаг - границы очага определялись по расположению плотных сетей волчьих магистральных троп, окружающих логова и дневки.
IV.Ь.2. Ассоциативные индексы у социальных видов животных и «Попарно ассоциативные социограммы»
Исторически индексы ассоциации разрабатывались для описания совместной встречаемости видов растений в пределах сообщества или места обитания (Dice, 1945). В течение последних 30 лет зоологи перенимали этот метод для оценки прочности ассоциаций между особями, в особенности, у социальных позвоночных животных, например, у львов (Schaller, 1972), у зебр (Penzhorn, 1984 цит. по Lenher, 1996), у дельфинов (Bejder et al.} 1997). В его основе лежит предположение о том, что физическая близость (например, членство в одной группировке) означает социальную привязанность, а количество времени, проведенного вместе, коррелирует с силой данной привязанности (Bejder et al., 1997).
Безусловно, в природе существует много примеров, кардинально не вписывающихся в это предположение. Ни у кого не вызывает сомнения, что длительность столкновения двух собак при агонистическом взаимодействии в зоне конфликтов между группами (Поярков, 1986) отнюдь не соответствует силе их привязанности друг к другу (согласно данному выше истолкованию). Подобный тип «привязанности» следовало бы трактовать иным способом. Очевидно, при сборе материала такого типа необходимо учесть поведенческую подоплеку развивающихся событий. Тем не менее, сам факт выбора того или другого партнера или допущения той или иной особи в «свою компанию» при миролюбивых взаимодействиях уже много говорит об отношениях между исследуемыми объектами. Таким образом, при определенных ограничениях индекс ассоциации может оказать хорошую помощь исследователю в поиске объективного численного показателя зарегистрированных привязанностей.
Ассоциативный индекс рассчитывается для каждой диады - пары особей - в исследуемой популяции или части популяции. В большинстве случаев диапазон индекса, по определению, находится между значением 0, т.е. данные две особи никогда не были встречены вместе, и 1 - данные две особи всегда встречались вместе. Отсюда, чем больше значение индекса, тем выше уровень ассоциации между данной парой индивидуумов.
Ленер (Lenher, 1996:507-510) приводит подробный обзор ассоциативных
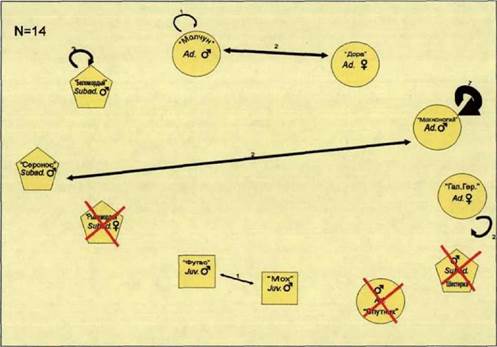
У.а. Состав групп
У.а.1 Воронежский заповедник
В общей сложности, в Воронежском заповеднике были учтены и идентифицированы 24 волка (см. таблицу 2), из них 18 самцов и 6 самок (соотношение полов: 3:1). Наши учеты полностью совпали с результатами годового учета, проводимого сотрудниками научного отдела Воронежского заповедника.
В таблице 8 отражен половой и возрастной состав восточной и западной групп волков, а также динамика их численности в период исследований.
Таблица 8. Волки «Восточной» (Е) и «Западной» ()Л/) стай. Половая и возрастная структура, а также изменение состава групп от зимы 1996-97 гг. к зиме 1998-99 гг.
ас/ - взрослый; эиЬас! - переярок; - прибылой; (+) - встречен в семейном участке стаи; (-) - не встречен на семейном участке. Красная линия указывает на особей, принимавших участие в размножении.
|
|
Кличка
(п=24)
Молчун Дора
Мохноногий
Галина Герм.
Спутник
Шестерка
Дядя Боря
Мучачо
Ял
Сеньорита__
Лука Чико
Беломордый
Серонос
Рыжемордая
Фугас
| Зима 1996-97 гг (п=17) Возраст ёЛ ^ здЬаё виЬас! виЬаё виЬас! - ^ Еще не родились |

| Зима 1997-98 гг |
| Зима Весна 1998-99 гг 1999 г =17) (п=14) |
| Щ ас! эиЬаё |
| ШУ Еще не родились |
| Еще не родились |
| Пол самец самка самец самка самец самец самец самец самец самка самка самец самец самец самка самец самец самец _| самка самец самец самец самец самец |
Мох
| | с* СО |
Лапастый
Борзайка
Троян Черный Шестерка II Лапась Борзай
Таблица 10. Половая и возрастная структура, а также изменение состава группы волков стаи «Зуевка» с марта 2000 г. по май 2001 г., национальный парк «Орловское полесье»
| 200 |
| I! |
| Пол (2,6:1) |
| Кличка (п=11) |

| (п=6) |
| Июнь (п =4) |
| Август (п=6) |
| Январь (п=6) |
| Май (п=5) |
| м о со |
| Атар | самец | ас! |
| Келта | самка | ас! |
| Партизан | самец | виЬаё |
| Риз | самец | эиЬаё |
| Дор | самец | виЬас! |
| Пир | самец | |
| Ал | самец | |
| Цид | самец |
| Лейка Алера Сар |
| самка самка самец |
аб - взрослый; эиЬаб - переярок; уиу - прибылой; (+) - встречен в семейном участке стаи; (-) - не встречен в семейном участке Красная линия указывает на особей, принимавших участие в размножении.
2000
Март
| 1 | - | - | -1 |
| .¡ЧУ | 1 . " | ||
| Еще не родились |
) - в январе 2001 г. зафиксированы две встречи группировок, состоящих из 11-ти волков, среди которых 5 животных индивидуально не определены.
(**) - по логике щенки в это времени уже родились, но мы их не встречали.
полесье» мы зарегистрировали и персонально идентифицировали 11 волков (см. Таблицу 5). Соотношение полов среди них составляло 2,66:1 (8 самцов и 3 самки). В таблице 10 приведены данные о половом и возрастном составе, а также динамике численности.
У.а.4. Заключение раздела
Исследованные группы довольно схожи по соотношению полов. Однако в соотношении возрастных категорий наблюдается заметный разброс. В Воронежском заповеднике у «Восточной стаи», в среднем по зимним данным, на каждого взрослого волка приходятся 0,81 переярка и 0,9 прибылых. У «Западной стаи» в этом же заповеднике соотношение возрастных категорий составляет 1:0,6:0,2. В заповеднике «Калужские засеки» — 1:1,8:1,6, и в национальном парке «Орловское полесье» —1:4,5:3.
Эти данные указывают на то, что, как стая «Зуевка» в «Орловском полесье», так и «Западная стая» Воронежского заповедника испытывают большой возрастной дисбаланс. У «Восточной стаи» Воронежского заповедника и стаи «Нагая» в «Калужских засеках» возрастные категории находятся в относительном равновесии.
\/.Ь. Семейный участок и его внутренняя структура
\/,Ь,1. Воронежский заповедник
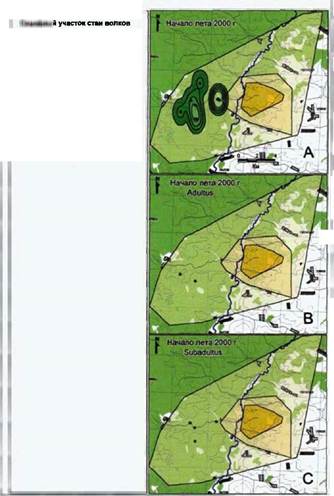
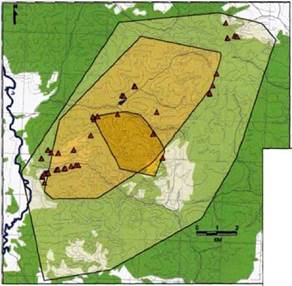
Семейный участок и его основные пространственные субъединицы (очаг, жизненное пространство и пространственная оболочка) восточной группы волков Воронежского заповедника в 1997 году отражены на рис. 13. Семейный участок восточной и западной групп в 1998-99 годах приводятся на рис. 14. Соотношения площадей пространственных субъединиц данных семейных участков приводятся в таблице 11.
В ] 998 году мы смогли структурировать семейный участок «Западной стаи» волков (что в 1997 году нам сделать не удалось из-за недостатка материала по этой группе), но его оценка не столь точна, как в случае «восточной группы». Во- первых, это связанно с тем, что часть семейного участка располагалась вне заповедной территории и была обследована в меньшей степени, во-вторых, некоторые компоненты очага западной группировки не были нами обнаружены, что затрудняет описание этой субъединицы. Также, неясны до конца и границы жизненного пространства, т.к. наблюдения охотничьего поведения этой стаи крайне немногочисленны.
Итак, перейдем к более подробному описанию полученных нами результатов по разным пространственным субъединицам «восточной стаи», в первую очередь в 1997 году.
Очаг
В нашем конкретном случае, одна из наиболее интересных особенностей очага состоит в асимметричности его расположения по отношению ко всему
|
Рис. 13. Семейный участок волков «Восточной стаи», Воронежский заповедник (1997 г.) |

| w J .' Логово |
| Ориентация на И" |
| ------ Волчьи тропы Выбитая площадка ц Бурелом с^Гэ Бугор |
Рис. 16. Первое логово. «Восточная стая» Воронежского заповедника (1997 г.) (верх.) схема расположения; (лев.) вид спереди.
окружающего древостоя доминирует сосна Pinus sylvestris с примесью дуба Quercus robur, хорошо развит подлесок, состоящий из клена Ar с es platanoides, орешника Corylus avellanarius и липы Tilus cordata. Логово использовалось до распускания листвы (со времени родов (приблизительно 06.04.97) по 26 апреля 1997), таким образом, «скрытность»
|
|

| Волчьи тропы Выбитая площадка Выворотня Бугор Л Поляна |
| Рис. 17. Второе логово. «Восточная стая» Воронежского заповедника (1997 г.) (верх.) окрестности логова ; (верх.лев.) вид спереди; (лев.) схема расположения. |
| Логово |
|
Рис. 21. (Лее.) Волчья магистральная тропа у третьей дневки, ведущая к Оброчному полю. (верх.) вид дневки. Воронежский заповедник (1997 г.) |

присутствуют черная ольха Alnus glutinosa и береза Betula sp., к востоку появляются дубы в сочетании с липами, в травяном ярусе доминирует осока волосистая Carex pilosa.
Последняя дневка представляет собой дневку третьей степени. Она имеет удлиненную форму, активно используется подросшими прибылыми (4-5 месяцев). Дневка довольно слабо структурирована: ее компоненты (игровые площадки, лежки, ВМТ) распределяются по достаточно большому участку, 300 м длиной и 50 м шириной. Оброчное поле (рис.21) дает прибылым относительную свободу движений (игры с преследованием, свойственные подросшим щенкам и т.п.) и возможность поупражняться в мышковании, обеспечивая одновременно возможность быстрого укрытия в случае опасности. Видимость на Оброчном поле - до одного километра.
В дополнение ко всему сказанному выше отметим, что последняя дневка и Оброчное поле как таковое представляют собой первую ступень в освоении щенками семейного участка. Освоение территории щенками происходит не постепенно, а ступенями. До того, как они заняли последнюю дневку, знакомство с территорией ограничивалось только частью очага и протекало очень медленно - сначала окрестности логов, потом маршрут между логовами и первой дневкой, затем последняя дневка. После этого темп освоения остальной части семейного
|
|
| Легенда Валки восточной группы Волки западной группы ф Зоны отдыха |
| Рис. 25. Движение волков вдоль границы семейного участка «Восточной стаи». Воронежский заповедник {1997 г,) |
Пространственная оболочка
Пространственная оболочка «восточной группы» волков простирается с самой восточной границы Воронежского заповедника до района линии электропередач, расположенной в его западной части. Пограничная линия между семейными участками двух существующих в Воронежском заповеднике стай в период с зимы 1996-97 года по зиму 1997-98 года подверглась изменениям, что было расценено нами как результат экспансии западной стаи на восток и формирование буферной зоны между группами. В 1998 и зимой 1998-99 - степень перекрывания семейных участков увеличилось, появился второй сектор на северо- востоке семейного участка «западной стаи». В указанный период семейные участки и восточной, и западной стай расширились в северном направлении.
На рисунках 25, 26, 27 изображены традиционные маршруты, используемые членами обеих стай, как в зоне контакта, так и на других границах пространственной оболочки на протяжении наших исследований.
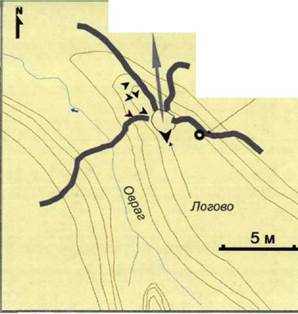
В травянистом ярусе вокруг логова доминирует осока волосисткая, Carex pilosa. Недалеко от него имеется валежник, что может служить волчице укрытием, но скрытое расположение логова и так делает его достаточно защищенным. Ко входу в логово со склона оврага спускается слабая тропа, еще одна протоптанная дорожка ведет от логова к ручью.
Это логово было занято в первой половине апреля.
Недалеко от первого логова (в 220 м) мы нашли еще одно, со сходными характеристиками, но недостроенное.
Второе логово расположено в 1200 м к юго-востоку от первого и в 800 м к северо-северо-востоку от единственной известной нам дневки. Оно построено также, предположительно, волчицей, под нависающим склоном оврага недалеко от его начала. Вход в логово ориентирован на север.
|
Рис. 31. Дневка на фаю очага. Южный участок «Калужских засек», апрель 2002 г. |

Существует предположение, согласно которому волчица могла перенести щенков из первого логова во второе и затем на дневку, хотя у нас отсутствует доказательства этого, так как с июля по октябрь на территории этой стаи исследования не проводились. О существовании дневки (рис. 29, 31) мы знаем по косвенным данным, таким как опрос сотрудников заповедника и жителей деревни Нагая, а также по тщательному рассмотрению post factum места, где дневка расположена. Дневка располагается между поляной и началом оврага, что предоставляет защиту волкам в случае опасности. Данная поляна расположена среди густых зарослей молодых берез и ив, видимость на ней не превышает 150-200 м. Поляна слабо дренирована, вода стоит под сухой травой, что делает невозможным бесшумное приближение к дневке.
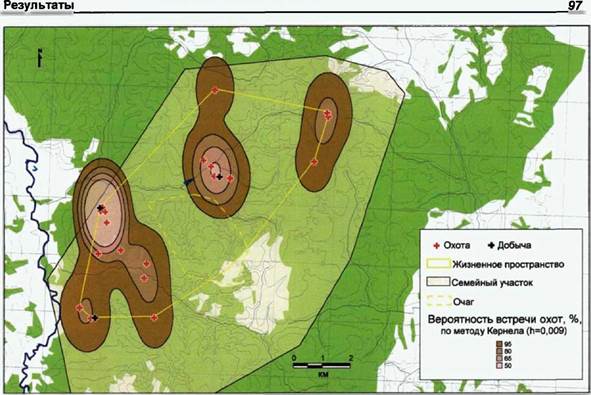
|
Рис. 32. Жизненное пространство волков стаи «Нагая» Южный участок заповедника «Калужских засек» (2000-02 гг.) Распределение охот (N=25). |
Жизненное пространство
|
Рис. 33. Расположение группировки семи волков. |
Как и в предыдущем случае, границы жизненного пространства были определены нами при помощи метода минимума по встречам мест охот группировок родительской пары с прибылыми. На рисунке 32 показаны границы жизненного пространства и распределение зарегистрированных нами мест охот. На этом же рисунке с помощью метода кернела мы решили проиллюстрировать фактические зоны наибольшей вероятности встреч охот.
| (§) Логово Дневка | Очаг ' Волчьи магистральные тропы |
У.Ь.З. Национальный парк «Орловское полесье»
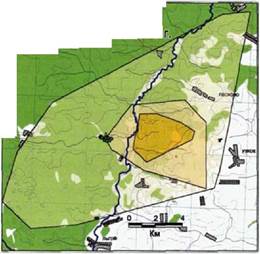
Семейный участок стаи «Зуевка» и расположение его очага, жизненного пространства и пространственной оболочки показано на рис. 35. Ниже следует более подробное
описание различных риг 36 0чаг волков стаи «зуеВка». Нац. парк «Орловское пространственных полесье» (2000-01 гг.)
| Ориентация на N |
| Лежка |
| Рис. 37. Первое логово. Нац. парк «Орловское полесье», май 2001 г. Серым цветом показаны ВМТ. |
субъединиц семейного участка стаи «Зуевка».
Очаг
Очаг расположен асимметрично по отношению к семейному участку, простирающемуся вдоль основной опушки лесного массива. Восточная граница очага примыкает вплотную к открытому полю и находится в 2,7 км от деревни Узкое (более 100 дворов) и в 1,7 км от деревни Сорокино (около 10 дворов). Лесной массив, находящийся на территории очага, состоит из различных древесных пород, рельеф варьирует от слабоволнистой равнины до пересеченной овражной системы. Русло ручья Песошня, пересекающее часть очага, занято
| Таблица 11. Общая площадь семейного участка и его пространственных субъединиц у стай волков: «Восточная» и «Западная» в Воронежском заповеднике (ВЗ) (1997 и 1998 гг.), «Нагая» в заповеднике «Калужские засеки» (КЗ) (2001-2002 гг.), «Зуевка» в национальном парке «Орловское полесье» (ОП) (2000-2001 гг.).
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
полесье» с другой. Первые две стаи практически лишены какого-либо давления со стороны человека. Последние две стаи находятся под воздействием антропогенного фактора, что может являться причиной небольшого размера жизненного пространства.
Относительная площадь пространственных оболочек семейных участков стай «Восточная» и «Нагая» с одной стороны, и «Западная» и «Зуевка» с другой, также сходна.
У всех исследованных стай мы наблюдали два способа устройства логова: расширение одного из отнорков барсучьего городка и самостоятельное строительство. В «Калужских засеках», где все известные нам барсучьи городки заняты самими барсуками, волки применяют второй способ. В «Орловском полесье» нам часто встречались нежилые барсучьи норы, с чем мы и связываем отсутствие самостоятельного строительства логова у волков. В Воронежском заповеднике два логова из трех волчица строила самостоятельно. К тому же, все они располагались на возвышении вдали от овражных систем.
Во всех рассмотренных нами случаях распределение элементов биологического сигнального поля не равномерно, логова находятся относительно близко друг от друга, но они отдалены от последней дневки. Только в «Орловском полесье» наличие отдаленного от других третьего логова нарушает предложенную схему.
К сожалению, невозможно провести сравнение характеристик дневок из-за
недостатка данных по «Калужским засекам» и «Орловскому полесью». Однако можно сказать, что во всех случаях последние дневки располагались вблизи границы очага, что подтверждает определение последней дневки как платформы, откуда начинается второй этап освоения пространства прибылыми. Здесь, забегая вперед, можно добавить, что данное освоение затрагивает не только физическое, но и социальное пространство.
У.с. Социальная организация и динамика использования пространства
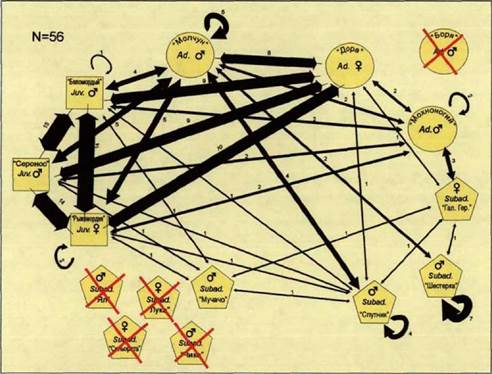
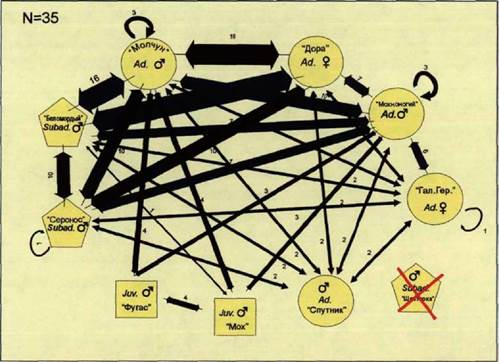
Ниже мы постарались дать описание социального контекста, на фоне которого протекали социальные процессы в стае волков на различных этапах жизненного цикла. Данные процессы, с нашей точки зрения, определяют характер использования пространства различными возрастными группами в течение года. Для более легкого восприятия данных для каждого периода жизненного цикла были построены попарно ассоциативные социограммы. Также далее приведены данные о распределении волков по территории семейного участка, полученные при помощи метода кернела. В заключении дана схема использования пространства волками в целом, т.е. с учетом всех годовых данных, а также суммарный результат кластерного анализа ассоциативных индексов волков каждой из стай.
\Лс.1. Воронежский заповедник
Зима 1996-97 гг. Эструс взрослой самки
Родительская пара, состоящая из самца Молчуна и самки Доры, находилась в очаге, и жизненном пространстве чаще всего в его западной части и в окрестностях Крутовского кордона (кв. 270). Их маршруты в этот период имели
циклический характер, они осуществлялись, в основном, против часовой стрелки, между очагом и кв. 241, к западу от железной дороги. Кроме того, пара появлялась в пойме р. Усманки, вдоль кв, 335, 314 и их окрестностей, где находились зоны отдыха (рис.22). Для пересечения железной дороги животные пользовались переходами вблизи Крутовского кордона (кв. 270), а также и немного севернее его (кв. 250).
Группировка, состоящая из взрослого самца Мохноногого, самца-переярка Спутника и самки-переярка Галины Герм., сохранялась на протяжении всего этого периода. Она была обнаружена в жизненном пространстве. Волки двигались по пойме от кв. 397 к кв. 331 вдоль ручья по стабильному маршруту.
Самец-переярок Шестерка часто встречался вблизи вышеуказанной группировки; но редко и ненадолго вступал в прямой контакт с нею.
Самец-переярок, названный нами Дядя Боря, появлялся на северных и южных границах очага, пересекая его по ВМТ в направлениях север-юг и юг-север (кв. 231), Его присутствие было обнаружено также вблизи Ледовских полян и Оброчного поля (соответственно, северная и южная границы очага). Он часто ходил по пойме в обоих направлениях от кв. 335 к кв. 194.
Группировка, которую составляют четыре из пята прибылых 1996 года рождения - ЬАучачо (с?), Яп{&\ Сеньорита (?) и Лука ($), использовала места охоты, расположенные на востоке от железной дороги в радиусе примерно двух километров от Крутовского кордона (270 кв). Недостающий в этой группировке прибылой Что, самый мелкий самец из выводка, никогда не наблюдался вместе со своими братьями. Он появлялся на ВМТ, проходящей с севера на юг между кв.192 и 193 и сопровождал своего отца, Молчуна. В начале февраля от группировки прибылых отделилась также волчица Лука, самая мелкая из двух самок помета. Она ненадолго присоединилась к родительской паре и сопровождала их по периферии очага. Таким образом, прибылые четко делятся на крупных, занимающих северозападную часть жизненного пространства западнее железной дороги, и мелких, находящихся вблизи западной границы очага.
Мы не располагаем данными относительно локализации членов западной группы зимой 1997 года.
На рисунке 41 показана попарно ассоциативная социограмма волков «Восточной стаи». Распределение различных возрастных категорий по
|
Рис. 45. Попарно ассоциативная социограмма волков «Восточной стаи». Осень 1997 г., Воронежский заповедник. |
|
|
| .. А<1, п=35 ЯиЬаа. п=38 п=46 |
| 2,8е/. |
34,2%
тпс
| □ 50,096 |

| 6,5% |
| 93,5%
|

п.
| 18,5%
|

0%
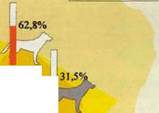
| Таблица 12. Временное распределение активности волков на Оброчном поле (Август- октябрь, 1997 г.) Воронежский заповедник.
|
сопровождении самки-переярка Галины Гермна западной границе жизненного пространства.
Присутствие самца-переярка Спутника было зафиксировано в кв.229 (около железной дороги напротив западной границы очага), куда он пришел со стороны р. Ивницы.
Самец-переярок Шестерка появлялся, в основном, один, западнее железной дороги (кв.227).
Взрослый самец Дядя Боря не был зарегистрирован; с большой вероятностью можно предположить, что он либо покинул семейный участок, либо погиб, так как в дальнейшем он также нами обнаружен не был.
Из пятерки переярков 1996 года рождения на семейном участке «Восточной группы» был зарегистрирован только самый крупный из них, самец Мучачо, встречавшийся в районе кв. 276 и 297.
Прибылые 1997 года рождения держались на последней дневке, время огг времени осуществляя спорадические заходы до Дедовских полян, то есть до мест, расположенных возле первой дневки.
В таблице 12 представлены данные визуальных наблюдений на Оброчном поле, с указанием времени появления волков на поле и времени их удаления. Из пятнадцати дней, во время которых проводились визуальные дежурства, в течение семи (46%) на поле можно было наблюдать от 1 до 5 волков.
Из членов «Западной группы» на границе между семейными участками
|
Рис. 49. Попарно ассоциативная социограмма волков «Восточной стаи». Весна-лето 1998 гг., Воронежский заповедник. |
Ad.Hr115 ! 5иЬас1. п=5 ,1иу. п=2
|
|
|
Рис. 53. Попарно ассоциативная социограмма волков «Восточной стаи». Зима 1998-99 гг., Воронежский заповедник. |
Ас|- «=69 I
-^Т БиЬас!. п=24
| ' 27,6% О«:
Ли ? 1 г » |

социограмме (рис. 61).
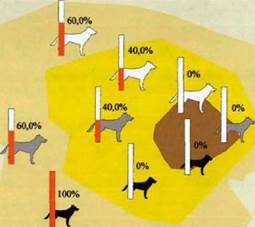
На рисунке 62 показана область встреч всех волков в этот период. Кернел встреч различных возрастных категории в семейном участке и их процентное соотношение по пространственным субъединицам показаны на рисунке 63. На этом рисунке видно, что волки всех возрастных категорий чаше всего отмечались в жизненном пространстве.
Весна-лето 2001 г.
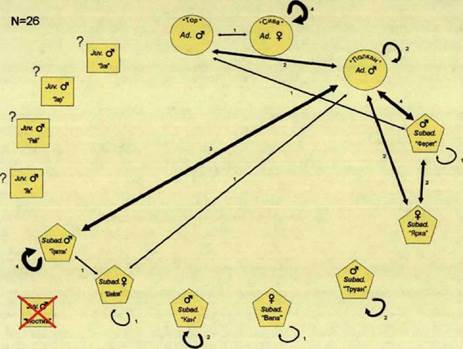
Родительская пара была встречена в очаге, как вместе, так и по отдельности. Самец Тор также был зарегистрирован у южной границы пространственной оболочки вместе со взрослым самцом Полканом.
Взрослый самец Полкан, встречался у северной границы пространственной оболочки, как один, так и в сопровождении самца-переярка Ферета и самки- переярка Ярки или самца-переярка Тритона и его сестры переярка Шейлы.
Кроме Ярки, все переярки встречались также и по отдельности. Тритон
|
|
|
Рис. 67. Попарно ассоциативная социограмма волков стаи «Нагая». Зима 2001-02 гг. (анэструс), «Калужские засеки». |
|
|
Начало зимы 2001-02 гг. Анэструс
Родительская пара вместе со взрослым самцом Полканом и четырьмя прибылыми 2001 года рождения (Зов, Зар, Рай и Як) образовали группировку, которая держалась, в основном, по северной половине жизненного пространства и пространственной оболочки,
сочетая в своих маршрутах
охотничке повеление г М Пространственное распределение волков стаи охотничье поведение с <<Нагая >к Зима 2001-02 гг. (анэструс); (N=110; Ь=0,012)
|
|
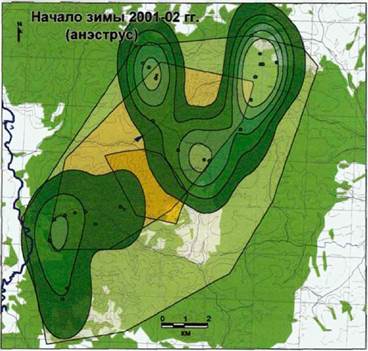
маркировочной активностью. (см- легенДУ Рис- 62) Взрослые самцы Тор и
Полкан встречались вместе на южной границе жизненного пространства и активно ее маркировали (см. Г\Лс12).
Самцы-переярки Труан, Кан и Мостик были встречены как на северной границе семейного участка, так и на южной. Несмотря на то, что проходы этой группировки, во многом, совпадали с маршрутами перемещений группировки родительской пары, взаимодействий между ними мы не обнаружили.
Переярки 2000 года рождения Тритон и Шейла отмечались вместе недалеко отд. Кирейково, на северной границы территории.
Самец-переярок Ферет встречался только по одиночке, там же, где и самец- переярок Тритон,
Самки-переярки Вела и Ярка в пределах семейного участка нами обнаружены не были. Последняя не была встречена и в дальнейшем.
В этот период, по сравнению с предыдущей зимой, взаимодействия между членами стаи приобрели асимметричный характер (рис. 67).
На рисунке 68 показана область встреч всех волков в этот период. Как взрослые, так и прибылые встречались и в жизненном пространстве, и в пространственной оболочке, в отличие от переярков, которые, не вступая во взаимодействие со взрослыми, держались в пространственной оболочке (рис. 69).
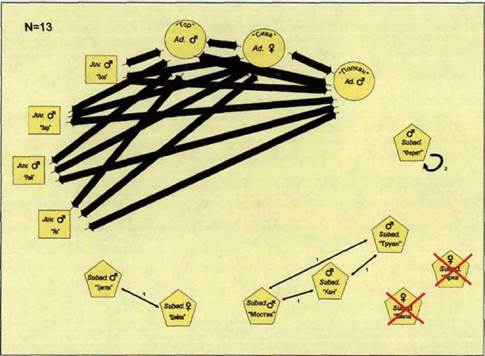
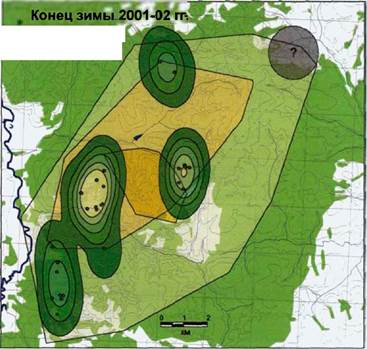
Конец зимы 2001-02 гг. Эструс
В середине января родительская пара все еще ходила вместе с четырьмя прибылыми и взрослым самцом Полканом. Это длилось до наступления эструса у взрослой самки Сивы (31.01.02г.).
В начале февраля родительская пара уже перемещалась по очагу отдельно от остальных волков. С ней продолжал оставаться только самый мелкий из прибылых 2001 года рождения Як. При троплении этой группировки выяснилось, что на маршруте Як часто отставал (5 раз за 3 км хода) от своих родителей и догонял их бегом, из чего можно предположить, что родительская пара (или один из взрослых) его отгоняла.
Остальные прибылые 2001 года рождения были обнаружены вместе со
эструса у взрослой самки все еще держалась вместе. В это же время нами был зафиксирован случай встречи родительской пары (вместе с Яком), с Полканом и остальными тремя прибылыми. Первые целеустремленно вышли из очага через его северовосточную границу, пересекли овраг и на
плакоре встретились со
втопой рпуппиппвкой Рмс- Т1- Пространственное распределение волков стаи второй группировкой, {<Нагдя , Зима 2001-02 гг. (эструс); (N=96; М=0,009) (см.
| Рис. 70. Попарно ассоциативная социограмма волков стаи «Нагая». Зима 2001-02 гг. (эструс), «Калужские засеки». |
| взрослым самцом Полканом. Таким образом, выяснилось, что часть группировки из семи зверей после начала |
| (эструс) |
пришедшей с севера. После легенду рис. 62)
встречи состав группировок не изменился. Тор, Сива и Як, описав большую петлю, вернулись к очагу почти по своим следам, Полкан, Зор, Зар и Раи ушли к востоку. На месте встречи никаких следов агрессивных взаимодействий обнаружено не было.
Переярки Труан, Кан, Вела и появившийся снова на территории семейного участка Мостик составляли группировку (в том же составе, что и зимой 2000-01 года), которую мы отмечали неоднократно в окрестностях деревни Ягодное, в южной части пространственной оболочки и жизненного пространства.
О судьбе переярков Ферета, Тритона и Шейлы мы не можем ничего сказать, поскольку в южной и центральной части семейного участка эти волки в указанный период встречены не были, а обследование северной части участка семьи из-за погодных условий в данный период не проводилось.
В конце зимы, как и в ее начале, взаимодействия между членами стаи «Нагая» имеют асимметричный характер (рис. 70). На рисунке 71 показано распределение волков по семейному участку в этот период. Встречи с взрослыми волками и прибылыми, преимущественно, регистрировались в жизненном пространстве, с переярками - в пространственной оболочке (рис.72).
Весна 2002 г. Щенки в I логове
Судя по собранному нами материалу, весной 2002 года состав стаи «Нагая» достаточно резко сократился. Несмотря на тщательное обследование территории семейного участка волков, мы не обнаружили семи переярков: Труана, Кана, Велу, Мостика, Зара, Зора и Яка (рис. 73). Скорее всего, они покинули семью, что представляется нам более вероятным, чем их гибель, учитывая количество животных. Переярки Труан, Кан, Вела и Мостик в 2000-01 гг. составляли сплоченную обособленную группировку, которая, по всей видимости, обрела самостоятельность.
Оставшийся переярок 2001 года рождения, Рай, был обнаружен нами у северной границы семейного участка, но с кем он там взаимодействовал и взаимодействовал ли вообще с кем-нибудь, остается для нас загадкой.
находится очень далеко от своих брата и сестры.
Мы бы хотели снова указать на судьбу щенка, тесно связанного с родительской парой по окончании первой зимы жизни: на судьбу Яка. Его взаимодействия с другими волками были очень слабы, в итоге, он очень рано покинул стаю.
\Лс.З. Национальный парк «Орловское полесье»
Весна 2000 г.
На территории стаи «Зуевка» в начале весны 2000 г не было найдено никаких следов или меток родительской пары, несмотря на обследования участка по снегу во всех пространственных субъединицах.
Четыре прибылых: три самца Ал, Пир, Цид и одна самка Лейка были найдены вместе с двумя самцами-переярками Партизаном и Ризом в окрестностях Михайловского кордона (рис. 79). Волки активно перемещались по заболоченным участкам возле кордона, а также между домами и сараями. В этот период других признаков, свидетельствующих о существовании волков, кроме их старых логов, мы не обнаружили.
Начало лета 2000 г.
|
|
| Локализация волков * 1 особь Семейный участок стаи волков | | Очаг I Жизненное пространство | [ Пространственная оболочка |
| Начало весны 2000 |
| Рис. 79. Пространственное распределение волков стаи «Зуевка». Весна 2000 гг., нац. парк «Орловское полесье». |
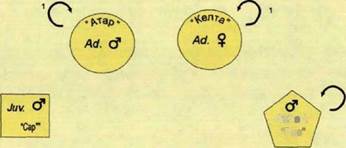
Взрослый самец Атар перемещался по западной границе семейного участка вдоль лесовозных дорог и по лесным просекам.
____________________________________________________________ Взрос-
|
|
|
|

N=6
|
|
| виЬай "Риз" |
Л/И ? Апера'
|
|
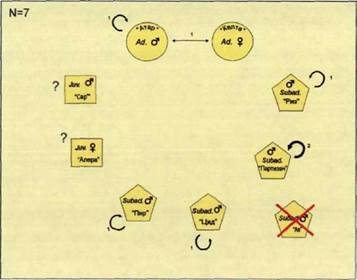
Рис. 80. Попарно ассоциативная социограмма волков стаи «Зуевка». Начало лета 2000 гг., нац. парк «Орловское полесье».
дельно от самца.
Самцы-Переярки Риз и Ал перемещались по просекам в долине ручья Шковка.
Взаимодействия между волками нами отмечены не были (рис.80). Все волки и их следы жизнедеятельности обнаружены западнее жизненного пространства, в пределах пространственной оболочки (рис.81).
Конец лета 2000 г.
| : Очаг 3 Жизненное пространство I [ Пространственная оболочка |
| • 1 особь Вероятность встречи волков, %, по мешду Карнела ■ к |
| Ш«5 □ 50 □35 |
| Рис. 81. Пространственное распределение волков ПеРеяРки были встречены как в стаи "Зу^. Начало лета 2000 гг. А-все особи очаге {Партизан, Риз), так и у (N=10^=0,008); В —взрослые; С —переярков. 4 ^ ' 3 западной границы семейного участка (Цид, Пир). Кроме указанной встречи взрослых волков, других взаимодействий между членами этой группы мы не зафиксировали (рис. 83). |
| Шковка. Взаимодействия между волками нами отмечены не были (рис.80). Все волки и их следы жизнедеятельности обнаружены западнее жизненного пространства, в пределах пространственной оболочки (рис.81). |
| Конец лета 2000 г. Взрослый самец Атар вместе со взрослой самки Келтой были обнаружены на левом берегу р. Вытебети южней д. Б. Михайловки (рис.82). |
|
|
| Зима 2000-01 гг. К зиме 2000-01 гг. социальная |
« 3 у е в к и » возросла,
Кластерный анализ с использованием ассоциативного индекса
средней взвешенности (Н\М)
Метод взвешенной средней связи 1-Пирсон "г"
Атар ас1.00 Келта ай.00 Алера |1иг.00 Сар ¡Ш1.00 Партизан зиЬасЮО Риг виЬас1.00 Дор зиЬасШ Пир ]ии.99 Лейка |ш.99 Ццд ЛЛ*.99 Ал |шг.99
20 30 40 50 60 70 80 90 100 110
(Расстояние связи/Макс, расстояние/100 Рис. 91. Анализ взаимодействий волков стаи «Зуевка» (2000-01 гг.).
социальный стержень или же он слабо выражен. Лишь некоторые волки (родительская пара, самцы-переярки Партизан и Риз) поддерживают более или менее устойчивые связи.
У.с.4. Заключение раздела
| 1 V |
| ||
| 1 |
| ||
| 1— |
| | | |
| 1 | |||
Результаты наших исследований показали, что волки используют различные части семейного участка с разной интенсивностью и также в разной степени взаимодействуют друг с другом. Распределение особей по семейному участку согласовано с определенной схемой, которая обусловлена, в первую очередь, особенностями биологии вида и повторяется с заданной периодичностью. Характер осуществления данной схемы зависит от различных факторов среды, как
физической, так и социальной, и может варьировать из года в год, что показано на примере волков «Воронежского заповедника».
Согласно нашим данным эта схема предполагает преимущественное использование жизненного пространства всеми членами стаи с осени по конец зимы. Весной все особи, не участвующие в размножении, как взрослые, так и переярки, рассредоточиваются по периферии семейного участка, освобождая тем самым лсизненное пространство. При этом взрослые животные, вступающие в размножение, занимают очаг. Самец совершает выходы в жизненное пространство для снабжения кормом самки и приплода. Как только щенки обретают способность к жизни на дневках, постепенно начинается процесс обратной консолидации всей стаи.
Нами было отмечена большая разница между весенними и зимними индексами группирования у стаи «Нагая» и «Зуевка», чем у «Восточной стаи». Вряд ли это связано с популяционными особенностями, мы больше склоняемся к другим причинам. На наш взгляд, такая разница в индексах характерна для молодых стай или стай в процессе формирования, хотя мы не исключаем и другие причины.
Анализ динамики социальных связей между особями одной и той же стаи показал, что, несмотря на разницу в сезонном характере группирования, связи между членами стаи поддерживаются в течение всего года. Эти связи осуществляются либо напрямую между всеми волками, либо посредством особей- звеньев. Роль связующих звеньев свойственна таким особям, как взрослый самец Мохноногий в «Восточной стае» и взрослый самец Полкан в стае «Нагая».
Однако утверждения, справедливые для стай «Восточная» в Воронежском заповеднике и «Нагая» в «Калужских засеках», совершенно не верны для стаи «Зуевка» в «Орловском полесье». Во первых, перемещения волков стаи «Зуевка» по территории семейного участка весьма хаотичны и характеризуется большой степенью иррегулярности, что, на наш взгляд, может являться прямым отражением особенностей социальной сферы этой группы. Мы считаем, что данная стая сформирована не до конца. Мы мало знаем о стадиях формирования стай, о возможных этапах их восстановления после разрушения существующих социальных связей путем физического уничтожения особей. Поэтому мы называем этих волков группой и допускаем при этом, что такое состояние постоянной
году резко увеличилось, что привело к увеличению длины буферной зоны.
Поскребы
Мечение поскребами изучалось многими исследователями (Peters, Mech, 1975, Grande del Brío, 1991 и др.), причем все они признают поскребы как запаховыми, так и визуальными метками
Рис. 98. Связь поскребов и (Bekkof, 1979), а также отмечают их автономность по других типов меток. (N=62).
отношению к экскрементам и мочевым меткам.
Полученный нами материал в Воронежском
заповеднике также подтверждает этот вывод (рис. 98).
Гранде дель Врио (Grande del Brío, 1991), предположил, что форма поскребов может свидетельствовать о возрасте зверя, оставившего их. Для подтверждения этого мы выделили три типа поскребов: сходящиеся, расходящиеся и параллельные, (рис.11).
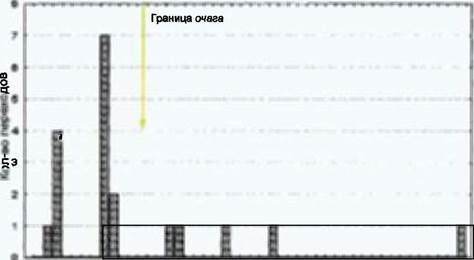
Из проанализированных нами в 1997 году 103 поскребов 45 (43,7%) оказались многосерийными (2-5 серий). Среди этих последних чаще всего встречаются двухсерийные (29; 50%), за ними следуют трех- (17; 29,3%), четырех- (10; 17,2%) и пятисерийные (2; 3,4%).
| смочеэыми |
| 36% |
| 'с экскрементами i 26% |
| отдельно 36% |
|
|

| Adultos ■ — |
| ___________________ Subadultus ■ Параллельные Q Расходящиеся | |
| Рис. 99. Соотношение различных типов поскребов в Воронежском заповеднике. |
Для 40 из 193 проанализированных нами поскребов нам удалось определить по следам, какой именно волк совершил данный поскреб. Соотношение возраста животного и типа поскреба представлено на рисунке 99, из которого видно, что большинство многосерийных поскребов являются сходящимися. На основании этой гистограммы можно прийти к выводу, что взрослые волки совершают, преимущественно, сходящиеся поскребы, тогда как у переярков, судя по всему, преобладают расходящиеся.
| Таблица 13. Характер перемещения волков, анализ количества расходов и резких поворотов (т - самец, 1 - самка).
I -I |
|
100 м интервал Рис. 106. Изменение количества переходов из колеи в колею при подходе к очагу. Воронежский заповедник, 1998 г. |
6. Маршрут группировки из «Западной стаи» во время вторжения на чужую территорию имеет явно более прямолинейный характер и почти не имеет ответвлений по сравнению с маршрутом той же группировки внутри границ ее семейного участка.
При перемещении волков по местам охоты наблюдалась тенденция к движению от возвышения к возвышению, на которых часто встречаются лежки копытных. Нередко взрослые волки, передвигаясь по ВМТ жизненного пространства, сворачивали с нее, чтобы обследовать возвышения, судя по всему, заранее им известные, так как животные двигались к ним напрямик. В том случае, если эти обследования не давали положительного результата, волки обычно возвращались на ВМТ и возобновляли прерванный путь.
В Воронежском заповеднике волки в зимнее время часто передвигались по замерзшим водоемам, так как глубина сугроба на льду значительно меньше, чем по берегам, что облегчает ход. Это явление, отмеченное и в других широтах (МесЬ, 1970), приобретает большее значение внутри очага и в жизненном пространстве, где реки и ручьи располагаются по периферии (в первом случае), или соединяют очаг с местами охоты (во втором случае).
Зимой, когда у нас была возможность проводить детальные тропления на большие расстояния, мы выяснили, что из 67,15 км, протропленных нами, 26%
|
|
| Семейный участок стаи волков I- . .Ючаг I_____ ¡Жизненное пространство I—_] Пространственная оболочка |
Результаты
|
|
| Мочевые метки волков Пространственное распределение А Зиыа |
| Семейный участок стаи волков I I Очаг | | Жизненное пространство | Пространственная оболочка |
| Рис. 107. Пространственное распределение мочевых меток на семейном участке волков стаи «Нагая». 2000-2002г., «Калужские засеки». |
У.е. Кормовая база и питание
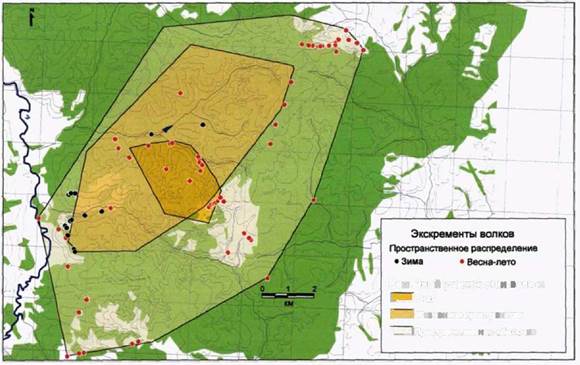
Нами было показано отсутствие прямых связей между распределением жертв и расположением субъединиц семейного участка волков в Воронежском заповеднике (Казьмин и др., 2000), где копытные отмечались во всех зонах семейного участка стаи в течение всего года. Нам казалось целесообразно выяснить распределение жертв и спектр питания волка и в других местах исследований.
Для этого мы провели анализ экскрементов, собранных в Воронежском заповеднике и в «Калужских засеках». Анализ питания волков «Орловского полесье» провести не удалось из-за малого количества материала.
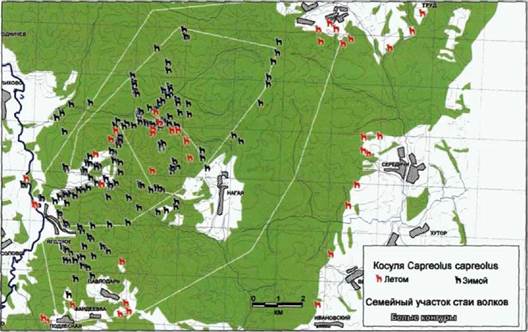
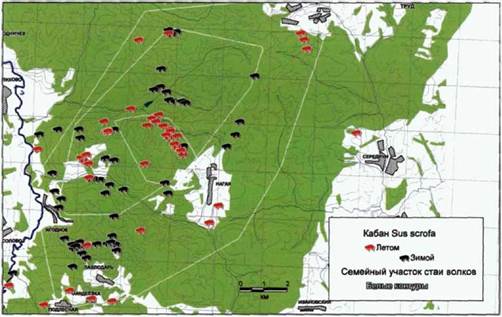
Распределение копытных на южном участке «Калужских засек» сильно зависит от времени года. Это особенно характерно для косули. На рисунках 116 и 117 показаны места регистрации встреч с кабанами и косулями в период 2000-2002 гг. В таблице 14 показано соотношение встреч/км маршрута для этих видов в
| Таблица 14. Соотношение встреч переходов трех видов копытных на км маршрута на южном участке заповедника «Калужские засеки», 2000-02 гг.
|
| зимний и весенний периоды. |
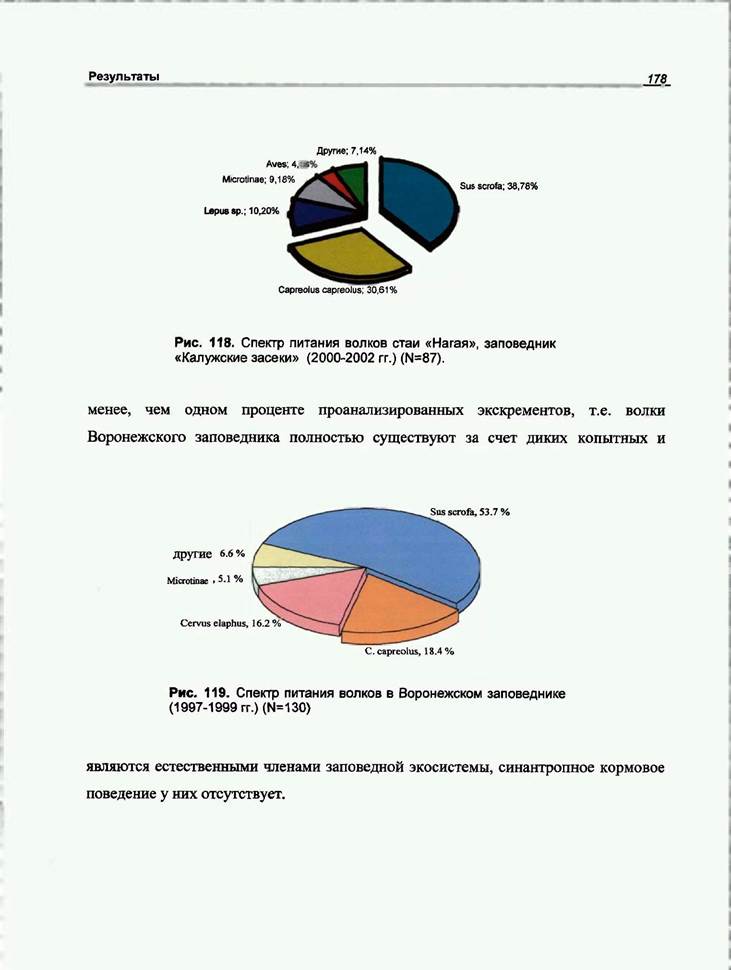
Для анализа питания волка в «Калужских засеках» было проанализировано 87 экскрементов. Результаты приведены на рисунке 118. Для анализа питания волка в «Воронежском заповеднике» проанализировано 123 экскремента. Основные результаты приведены на рисунке 119.
Проведенный нами анализ показал, что у стаи «Нагая» кабан и косуля равно представлены в питании. Это дает возможность предположить наличие сезонной смены доминирующего вида жертвы. К сожалению, небольшая выборка по сезонам не позволяет провести отдельный анализ весеннего и зимнего питания. Также нам представляется интересным значительное присутствие в рационе небольших видов жертв, таких, как заяц и мышевидные грызуны.
В Воронежском заповеднике кабан является основной жертвой волков
|
Рис. 116. Распределение косуль на семейном участке волков стаи «Нагая», (20002002 гг.) (по количеству встреч на маршруте). |
|
Рис. 117. Распределение кабанов на семейном участке волков стаи «Нагая», (20002002 гг.) (по количеству встреч на маршруте). |
|
|
локализации в неделе (Okarma et al., 1998). Таким образом, изучение параметров использования пространства волками методом радиослежения не менее трудоемкое занятие, чем выполнение этих же задач с использованием применяемого нами методологического комплекса. Согласно нашим данным, годовой семейный участок, определенный методом минимального полигона, используется очень избирательно, всеми членами стаи, что, в первую очередь, зависит от характера социального контекста. На наш взгляд, необходимо учитывать данный контекст при интерпретации данных, полученных методом радиослежения. В литературе широко практикуется определение индивидуальных участков в течение недлительного периода времени и определение «семейных» участков нескольких меченых животных в стае (н.п. Boitani, 1982; Fuller, 1989). На наш взгляд, эти данные совершенно не сопоставимы, что хорошо видно по нашему материалу. Существует определенная путаница между понятием «степень использования участка» (как правило, годового) с самым понятием индивидуального участка. С учетом того, что волк является высоко социальным видом, с нашей точки зрения правильный подход к изучению его пространственной организации базируется на определении семейного участка как участка обитания всей группы, а затем на характере использования этого участка каждым из ее членом, что и было показано нами. В Польше также была показана разная степень использования годового семейного участка каждым членом группы, но без учета социальных связей (Jedrzewski et al., 1998).
На примере волков Воронежского заповедника мы показали, что изменения в социальной сфере, такие, как отсутствие щенков у родительской пары или изменение обычной картины социальных связей, могут отражаться на размере семейного участка. Этот влияние подобных факторов на размер участка также было отмечено ранее (Okarma et al., 1998). Площадь изученных нами семейных участков соответствует показателям, полученным в других местах ареала вида как в Евразии (Калецкая, 1973;Vila et al., 1990; Okarma et al., 1998), так и в .Северной Америке (Fuller, et al., 1992). Согласно Г. Окарме с соав.(Окагта et al., 1998), у польских беловежских волков размер участка варьировал в пределах 141-168 км2 (мая- сентябрь) и 90-271 км2 (октябрь-апрель). В нашем случае семейные участки не претерпели изменения в течение одного жизненного цикла, но изменения были отмечены между годами. Отсутствие заметных изменений в размере участка между сезонами (между весной и зимой) объясняется перераспределением членов стаи. Данное перераспределение таково, что всегда одна или несколько особей отмечаются на периферии участка, от чего и зависит расположение крайних локализаций. Мы считаем нецелесообразным выявлять изменения размера семейного участка в приделах года. Во первых, если согласиться с Окарма с соав. (там же) о том, что необходимо 9-12 месяцев для получения верных данных о размере участков с помощью радиослежения, тогда говорить о каких либо сезонных изменениях этого параметра просто неадекватно, поскольку эти данные будут неполноценными. С другой стороны, гораздо информативно получение данных о степени использования того же самого годового участка.
Площадь семейного участка волков, живущих в условиях более бедной кормовой базы (стая «Зуевка» в «Орловском полесье») больше, чем у волков в условиях более богатой кормовой базы (стая «Нагая» в «Калужских засеках»). Эта же закономерность была прослежена в Квебеке (Messier, 1985). В нашем конкретном случае не стоит упускать из внимания и такой важный фактор, влияющий на размер участка, как стабильность. В Швеции (Bjarvall, Isakson, 1982) и Португалии (Pereira et al., 1985) было показано, что, чем менее стабильна группа волков, тем больше участок, которой она занимает. Авторы приводили пример групп, находящихся в стадии расселения, как высоко нестабильных, занимающих большой участок. На наш взгляд, этот фактор, наряду с бедность кормовой базы, может оказывать влияние на волков национального парка «Орловского полесье».
Несмотря на то, что в 1998 году у волков «восточной стаи» Воронежского заповедника семейный участок увеличился, процентное соотношение пространственных субъединиц практически не изменилось. Площадь очага, в процентном соотношении, у всех представленных стай очень сходна (около 6% от размера семейного участка), что говорит о сходных характеристиках расположения элементов биологического сигнального поля у семейных групп волка. Данные других авторов подтверждают этот явление. Так, в Польше, согласно данным радиослежения, размножающаяся самка в мае использует около 5% годового семейного участка (Jedrzejewski et al., 2001).
Относительная площадь второй территориальной субъединицы — жизненного пространства сходна у тех стай, которые живут в похожих условиях существования. Это относится к стаям «Восточной» в ВГЗ и «Нагая» в ЗКЗ.
Площадь перемещения и отдаления от дневки зависит от наличия щенков в стае. Так, было показано, что зимнее перемещение волков без щенков меньше, чем с щеками (Jedrzejewski et al., 2001; Mech et al., 1998). Отсюда правильно предполагать, что преимущественное совместное использование жизненного пространства всеми членами стаи будет имеет место с осени по конец зимы. Это же показали наши результаты. Весной все особи, не участвующие в размножении, как взрослые, так и переярки, рассредоточиваются по периферии семейного участка, освобождая тем самым жизненное пространство. Этот феномен неоднократно отмечался раньше (Кудактин, 1981) (хотя и интерпретировался как временное отщепление переярков от стаи). При этом взрослые животные, вступающие в размножение, занимают очаг. Самец совершает выходы в жизненное пространство для снабжения кормом самки и приплода. Как только щенки обретают способность к жизни на дневках, постепенно начинается процесс консолидации всей стаи. Анализ этой схемы объясняет, почему многие авторы считают что, «весенний» семейный участок меньше «зимнего» (Mech, 1970; Кудактин, 1981; Ballard et al., 1987), ведь для определения первого они используют площадь, занятую родительской парой (что в нашей работе практически совпадает с жизненным пространством), и не учитывают переярков-членов стаи, находящихся по периферии (в нашей работе пространственная оболочка). Исходя из этого предположения, в качестве сравнения можно было бы утверждать, что площадь весеннего участка примерно равна площади зимнего участка, который лишен периферийной субъединицы - пространственной оболочки, т.е. на 60% меньше (по нашим данным), что не противоречит данным польских авторов (весенний участок на 68% меньше, чем зимний) (Okarrna, 1998). Эта аналогия позволяет сравнивать наш материал с данными, полученными другими авторами.
Наши данные оправдали выделение пространственных субъединиц в рамках предложенного нами фунционально-типологического подхода к пространственной организации семейных групп волков. В этой работе мы показали впервые, что концепция трех субъединиц, являющаяся теоретической моделью, имеет практическое значение в исследовании волков, так как она отражает реальные процессы, протекающие в обществе хищника.
Более тридцати лет тому назад Адаме и Дейвис (Adams, Davies, 1967) подчеркнули в своей работе, что участки обитания млекопитающих используются хозяевами неравномерно, им свойственна внутренняя структуризация. Безусловно, наш материал вполне подтверждает это высказывание. Уже с самого начала возникновения объективных исследований экологии волка авторы почувствовали необходимость выделения центральной области территории волка, где проистекает самая скрытая часть его существования (Mûrie, 1944; Mech, 1967). Ключевым параметром для выделения такой области стало наличие логова. На самых ранних этапах выращивания щенков родительская пара привязана к логову. Эта привязанность определяет паттерн использования ими семейного участка в весенний период (Jedrzejewski et al., 2001; Mech, 1999). В литературе существует термин, обозначающий ту область участка, которая соответствует наиболее посещаемой области участка — ядерная зона (core area) (Kaufmman, 1962). Ядерная зона — площадь, где вероятность встречи с данным животным наибольшая — определяется как область, включающая 50% центральных локализаций животного. Мы можем утверждать, что очаг -это ядерная зона размножающейся самки в весенне-летний период. В литературе термин ядерная зона широко применяется для обозначения части пространства, где активность волков развивается более интенсивно в определенный период. Подобным образом говорят о ядерной зоне конкретной группировки переярков в зимний период и т.д. (н.п. Okarma et al., 1998). Из-за широкого применения этого термина в разных ситуациях мы вынуждены использовать термин очаг, предложенный ранее Р. Гранде дель Брио.
В отечественной литературе В.П. Болотовым (1986) был предложен термин выводковый участок, который автор определял как область в 20 км2 вокруг дневки (центр акустической активности в осенний период). На наш взгляд, для конкретного примера Центрального лесного заповедника, где работал В.П. Бологое , такое предложение очень удачно. Но заранее фиксированная площадь не будет соответствовать реальной зоне действия волков всего ареала, поскольку те 20 км2, которые являются 5% семейного участка волков в ЦЛГЗ, для участков других волков, например, воронежских, будут соответствовать примерно 15% семейного участка, что сильно выходит за рамки очага в нашем определении. К тому же дневки, как показано в настоящей работе, располагаются на границе очага, а не в его центре.
Одной из важных характеристик очага является его консерватизм. Консерватизм очага, на наш взгляд, является следствием консервативности волка в использовании логов и дневок (Сабанеев, 1877; Зворыкин, 1930; Mûrie, 1944; Mech, 1970; Рябов, 1993). Волки Воронежского заповедника, жившие там до 50-х гг., уже использовали район очага нынешней «Восточной стаи» (Рябов и др., 1992). Нам кажется, что ключ к пониманию такого консерватизма лежит в концепции биологического сигнального поля (Наумов, 1973). Особенно важно то, что очаг не имеет временного сезонного характера. В очаге выделяется собственная структура наиболее значимых в социальном плане элементов биологического сигнального поля. Принципиально важная роль очага была показана нами на примере волков Воронежского заповедника. В 1998 году родительская пара не размножалась, тем не менее, щенки другой пары взрослых волков, родившиеся на периферии семейного участка, уже к середине августа оказались в традиционном для семьи очаге. Это, на наш взгляд, подчеркивает то, что очаг является осевой, наиболее консервативной частью территории группы. Не менее важно и то, что очаг, скорее всего, очень привлекателен для всех членов группы. В этом смысле его можно рассматривать в контексте концепции аттракторов, предложенной М.Е. Гольманом и Е.П. Крученковой (1999), как организатор поведения членов группы в масштабах жизненного цикла волка.
Если термин очаг под тем или другим названием в литературе применялся ранее, понятие жизненного пространства кажется нам принципиально значимым оригинальным предложением, впервые появившимся в пионерских работах Р. Гранде дель Врио (Grande del Brio, 1988). Его выделение совершенно правомерно и подтверждается режимом использования семейного участка и закономерностями запахового мечения (см. результаты мечения экскрементами). В науке обсуждался пространственный градиент запаховой маркировки (Peters, Mech, 1975; Rothman, Mech, 1979; Сизонов, 1981). В этих работах о градиенте говорится как о величине с постоянным направлением, тогда как на самом деле его направление и показатель меняются в зависимости от сезона жизненного цикла. Именно динамика мечения приводит к тому, что мнения различных авторов подчас расходятся диаметрально. Так, Питере и Мич (Peters, Mech, 1975), а также Сизонов (1981) считают, что концентрация возрастает к границе территории, тогда как Никольский и Фроммольт (1989) констатируют, что дневки, т.е. центральные части семейного участка метятся наиболее интенсивно. Однако, как показали наши данные, никакого противоречия в этом нет, так как наблюдения первых относятся к зимнему сезону, т.е. к периоду периферической активности группы, а наблюдения вторых относятся к летному сезону, периоду центральной активности. Опять повторим, что пространственные субъединицы не только влияют на закономерности различных форм коммуникации, но и по-разному используются в течение жизненного цикла. С нашей точки зрения, динамика использования жизненного пространства в течение цикла имеет под собой важный биологический смысл. Он заключается в том, что весной, в отличие от зимнего периода, переярки не часто посещают жизненное пространство. Таким образом нагрузка на потенциальных жертв снижается. Что, в своей очередь, облегчает задачу выкармливания щенков родительской паре в конце весны — начале лета. В этот период самка строго не допускает присутствие переярков в районе логова и проявляет доминантное поведение по отношению к своему самцу (Mech et al., 1998; Mech, 1999)
В отличие от первых двух, более центральных субъединиц, третья пространственная субъединица, названая нами пространственной оболочкой, является менее стабильной (более динамичной) и в действительности выполняет защитную функцию. Эта закономерность очень хорошо видна при сравнении данных, полученных в Воронежском заповеднике в 1997 и 1998 годах. Так, на примере пространственной организации восточной группы волков в Воронежском заповеднике видно, что образовавшаяся буферная зона заняла часть пространственной оболочки из-за экспансии западной стаи в 1998 году. Характерно, что жизненное пространство не подверглось этой экспансии, которая была остановлена на его границе (зона наиболее интенсивных маркировок). Однако динамизм оболочки зависит не только и даже не столько от ее защитных функций, сколько от психологических характеристик волков, занимающих ее, так как пространственная оболочка является зоной преимущественной активности переярков и именно переярки, в отличии от взрослых, демонстрируют менее стереотипное и более динамичное поведение, как уже отмечал Р.Гранде дель Брио (Grande del Brío, 1991).
При изучении социальных взаимодействий главным орудием исследователя является метод визуальных наблюдений, поскольку этот метод позволяет наиболее адекватно зафиксировать процессуальносгь социальных событий (Овсяников, 1986). Однако применение этого метода затруднительно и иногда невозможно, особенно в условиях леса. К тому же, даже в открытых ландшафтах находиться в постоянным визуальных контактом с объектом представляет собой очень трудную задачу, в особенности, когда объектов несколько и они широко перемещаются (МесИ, 1995а). Для получения данных о социальной организации волков в настоящей работе мы прибегали к косвенным методам, позволяющим, однако, индивидуально идентифицировать изученных особей с большой точностью. Безусловно, наша методика не позволяла получить прямых данных о персональных отношениях между животными, но зато обеспечивала большую точность в определении наличия социальных контактов (социальных связей). Этот главный критерий, использованный нами для вычисления ассоциативных индексов, позволяет экстраполировать полученные результатов на более высокий уровень значимости. Такое утверждение основано на том, что волк, будучи высоко социальным видом и имеющий возможность ограничивать количество и характер своих социальных контактов, будет ассоциироваться с другими особями в соответствии со своей внутренней мотивацией. Мы считаем, что данная интерпретация адекватна стоящим перед нами задачам. Первое, что позволило нам судить о социальной структуре — это характер группирования. Отмеченная нами разница между весенними и зимними индексами группирования у стаи «Нагая» и «Зуевка», с одной стороны, и у «Восточной стаи», с другой вряд ли связана с популяционными особенностями, мы больше склоняемся к другим причинам. На наш взгляд, такая разница в индексах характерна для молодых стай или стай в процессе формирования, хотя мы не исключаем и другие причины (Калецкая, Филонов, 1987). Анализ динамики взаимодействий между особями одной и той же стаи показал, что, несмотря на разницу в сезонном характере группирования, связи между членами стаи поддерживаются в течение всего года. Эти связи осуществляются либо напрямую между всеми волками, либо посредством особей- звеньев. Роль связующих звеньев свойственна таким особям, как не вступающие в размножение взрослые самцы стай «Восточной» и «Нагая».
Однако утверждения, справедливые для стай «Восточная» в ВГЗ и «Нагая» в ЗКЗ совершенно не верны для стаи «Зуевка» в НПОП. Во первых, перемещения волков стаи «Зуевка» по территории семейного участка весьма хаотичны и характеризуются большой степенью иррегулярности, что, на наш взгляд, может являться прямым отражением особенностей социальной сферы этой группы. Мы считаем, что данная стая сформирована не до конца. Мы мало знаем о стадиях формирования стай, о возможных этапах их восстановления после разрушения существующих социальных связей путем физического уничтожения особей. Поэтому мы называем этих волков группой и допускаем при этом, что такое состояние постоянной социальной неопределенности может быть характерно для стай, находящихся под прессом со стороны человека, что, в свою очередь, может являться адаптивной поведенческой стратегией в изменчивой среде.
Распределение меток у стай «Восточная» и «Нагая» соответствует схеме динамики распределения особей в пространстве. Запаховые поля распределены вблизи границ очага, буферной зоны и пространственной оболочки, при приближении к той или иной границе интенсивность запаховых полей возрастает. При отсутствии буферной зоны запаховые поля располагаются у границы жизненного пространства.
Для молодых групп или групп в процессе формирования характерно слабое использование поскребов. Это особенно хорошо проиллюстрировано для стаи «Зуевка», но также характерно и для стаи «Нагая» в «Калужских засеках», в отличие от стаи «Восточная» Воронежского заповедника.
В процессе социализации волков можно выделить три основных периода. Первый период протекает в обществе родителей и собратъев-прибылых. Длится этот период от самого рождения до момента появления переярков на дневке. Наш материал недостаточен для того, чтобы обрисовать временные границы этого периода, так как мы не смогли установить, в каком возрасте щенков начинается подтягивание переярков к дневке. Следом за A.A. Никольским и К.-Х. Фроммольтом (1989) мы полагаем, что инициатором такого процесса может быть взрослая самка, которая является главным фактором консолидации стаи. Роль самки как регулятора сплоченности группы находит подтверждение и в наших данных по акустической активности волков Воронежского заповедника. Одновременно с первым периодом социализации происходит и первый этап освоения пространства.
Второй период социализация совпадает со вторым периодом освоения территории. Нами было установлено, что прибылые «Восточной стаи» 1997 года рождения начали освоение территории в возрасте шести месяцев. Согласно Я.К. Бадридзе (1987), в этом же возрасте, при экспериментальных условиях, наблюдается максимальное количество правильных обходов непрозрачных предметов при преследовании добычи или сверстников. Последнее указывает на установление высокого уровня рассудочной деятельности.
Во время второго периода социализации и освоения пространства прибылые знакомятся с территорией семьи и с другими особями группы помимо родительской пары, перемещаясь с ними по территории (Mûrie, 1944; Pimlott, 1975; Бологов, 1983). Влияние доминантного самца на прибылых было показано нами на примере первого взрослого самца в стае «Нагая» в «Калужских засеках» и на примере первого взрослого самца в «Восточной стае» Воронежского заповедника. Если раньше мы рассматривали главную взрослую самку как инициатора консолидации в стае, на этом этапе мы рассматриваем главного взрослого самца как инициатора начала процесса освоения территории щенками, для чего он использует механизмы управления внимания щенков, направляя их от дневки к периферии. Нами было показано, что одиночный вой взрослого самца при приближении к дневке был зафиксирован преимущественно на юго-западе от нее - при наступлении второго этапа освоения территории прибылыми последние начинали освоение жизненного пространства именно с этого места и в этом направлении.
Третий период социализации совпадает с наступлением эструса у самки и пребыванием родительской пары в очаге. Прибылые в первый раз в жизни начинают существовать вне прямой зависимости от родительской пары и распределяются по периферии участка. На этом этапе установление персональных связей между прибылыми и переярками кажется нам очень существенным фактором, определяющим успех социализации прибылых. На нашем материале показано, что те прибылые, которые стремятся не покидать общество родительской пары, не переходят на третий этап и далее практически не взаимодействуют с членами группы. Эти животные, в дальнейшем, первыми покидают родительскую группу.
Анализ динамики взаимодействий между особями одной и той же стаи показал, что связи между членами стаи поддерживаются в течение всего года. Эти связи осуществляются либо напрямую между всеми волками, либо посредством особей-звеньев. Роль связующих звеньев свойственна таким волкам, как второй взрослый самец в «Восточной стае» Воронежского заповедника и второй взрослый
самец в стае «Нагая» «Калужских засек».
Перемещения волков стаи «Зуевка» по территории семейного участка весьма хаотичны и характеризуется большой степенью иррегулярности, что, на наш взгляд, может являться прямым отражением особенностей социальной сферы этой группы, также как и неупорядоченности элементов ее биологического сигнального поля. Мы считаем, что данная стая сформирована не до конца. Мы мало знаем о стадиях формирования стай, о возможных этапах их восстановления после разрушения существующих социальных связей при физическом уничтожении особей. Поэтому мы допускаем, что такое состояние постоянной социальной неопределенности может быть характерно для стай, находящихся под прессом со стороны человека, что, в свою очередь, может являться адаптивной поведенческой стратегией в изменчивой среде.
Нами был зафиксирован интересный факт приуроченности последней дневки к границе очага. Мы придаем этому феномену большое значение, так как это может облегчать приток переярков в период максимального сплочения стаи. Вторым важным следствием такого выноса может являться то, что при самостоятельной активности прибылые тяготеют к центру очага, и это затрудняет освоение следующей территориальной субъединицы — жизненного пространства. Очень интересно, что освоение жизненного пространства начинается с зоны акустической преференции доминирующего самца, как было указано ранее. Можно предположить, что его акустическая активность облегчала прибылым начало освоения жизненного пространства. Отметим, что прибылые 1998 года рождения оказались именно в этой части и держались там дольше, чем в других местах.
Следует отметить, что характер хода, выявляемый при троплении, может выступать как индикатор эмоционально-мотивационного состояния животных. Так, по ряду формальных критериев можно выделить охотничьи, территориально- маркировочные, поисковые и другие маршруты. Это выгодно отличает метод наземного тропления от телеметрии, с помощью которой удается выявить, активен или не активен зверь, но нельзя сказать ничего о его мотивационном состоянии. Особенно ценные результаты можно получить, сочетая метод тропления с методом кинологической идентификации (Соколов, Сулимов, Крутова, 1990). Поскольку каждая особь имеет свой неповторимый индивидуальный запах и этот запах может быть воспринят и опознан собакой, каждый индивид, оставляя запаховую метку при акте маркировки, предоставляет исследователям свою «визитную карточку». В определенном смысле это может быть приравнено к акту радиопеленга, т.к. позволяет опознать индивида в конкретном месте. Кроме того, метод кинологической идентификации имеет безусловное преимущество перед радиослежением при изучении закономерностей запахового мечения.
Одним из важнейших аспектов поведения волка в том, что касается прокладывания маршрутов, является характер патрулирующего хода, особенно в буферных зонах. Д. Мич (Mech, 1994) показал, что в этих зонах часто происходят столкновения между группировками, которые могут привести к смерти части животных, в том числе доминантов. В нашем случае высокая частота патрулирования членами обеих групп волков в Воронежском заповеднике, а также напряженность, нашедшая свое выражение в интенсивности мечения, могли позволить предположить, по аналогии с материалом Мича, высокую возможность столкновения. Однако нами не было зарегистрировано ни одного подобного случая. Параллелизм, замеченный в маршрутах обеих групп, при более детализированном изучении показал, что проходы волков, принадлежащих к разным группам, никогда не происходят одновременно. Было замечено, что звери западной группы всегда перемещаются по границе по часовой стрелке, тогда как члены восточной группировки передвигаются против часовой стрелки. Частота проходов первых явно выше, нежели частота проходов последних, это может объясняться тем, что «Западная группа» имеет меньший семейный участок и более мотивированна в расширении участка и его охране. Кроме того, мы считаем, что западная группа более молодая и окончательно не устоявшаяся, т.е. социальная позиция членов группы не определена окончательно. Все приведенные факты показывают, что волки избегают потенциального конфликта, не отказываясь от прохождения через буферную зону, как утверждают другие авторы в применении к другим широтам (Mech, 1977; Grande del Brío, 1991; Mech, 1994), а проходят по ее территории в противофазе. Все это позволяет предположить, что речь идет о своего рода семиотическом конфликте. Явное отличие характера маршрута вторжения от маршрута обычного патрулирования у волков «Западной группы» наводит на мысль о том, что завоевание чужого пространства происходит поэтапно, как результат включения дополнительных заходов на незнакомую территорию. Нанесение элементов биологического сигнального поля играет решающую роль в модификации структуры территории и в освоении нового пространства.
Освоение нового пространства очень часто сочетается с формированием новой группы (Rothman, Mech, 1979), когда вновь образовавшаяся стая осваивает новую территорию. Однако существуют и другие пути расширения и сохранения существующей структуры популяции. Первый из них условно может быть назван приватизацией части старого семейного участка определенной группировкой внутри социальной группы. Это влечет дробление семейного участка и социальной группы на две или более частей, как это наблюдалось на Айл Роял (Peterson, 1977) и в ряде других районов. Однако этот процесс влечет за собой определенные отрицательные последствия для обеих групп. «Материнская» группа теряет устойчивость и пространство, т.к. роль покинувших группу зверей остается вакантной. Отделившаяся «дочерняя» группа вынуждена формировать биологическое сигнальное поле заново, существуя при этом на ограниченной территории и используя весьма ограниченные ресурсы, что, как правило, препятствует размножению новой группы в первый год после отщепления. Еще одной опасностью для этой группы является возможность возникновения конфликтов с прежними сородичами по стае.
Другой формой расширения и сохранения существующей структуры популяции можно считать эффект репликации. В стае, по мнению большинства специалистов, размножается только одна самка, и именно она проявляет агрессивное поведение в присутствии других самок (Zimen, 1976b). Однако в литературе много данных, подтверждающих существование нескольких выводков в одной стае, как у волка, так и у других видов. За двадцать лет изучения волка на Пиренейском полуострове Р.Гранде дель Брио (Grande del Brío, 1991: 101) работал более, чем с одиннадцатью стаями на разных ступенях развития. В одной из этих стай было отмечено появление на одном и том же семейном участке трех выводков, которые распределялись следующим образом: первый, принадлежавший доминантной паре и состоявший из пяти щенков, занимал область очага,; второй и третий выводки, из трех и двух щенков, соответственно, занимали разные части пространственной оболочки и принадлежали взрослым самкам-членам стаи. Это редкое явление наблюдалось и в других широтах. Подобная ситуация (Zvorovsky,
Pechenyuk, 1995) была описана и в Хоперском заповеднике. Во всех вышеуказанных случаях было отмечено, что периферийные пары после исчезновения доминантных пар (естественная или насильственная смерть) копировали структуру очага со всеми составляющими биологического сигнального поля. В Калининской и Кировской областях зафиксированы три случая, когда приплод разных самок располагался менее чем в 2 км друг от друга (Павлов, 1990). В Алтайском крае также регистрировалось наличие двух самок со щенками на одном «гнездовом участке» (Бондарев, 1979). А. Кудактин располагает сведениями о размножении на Кавказе более одной самки в одной и той же группе (1998).
Существуют также случаи «сложной семьи», согласно определению Пояркова и Покровского (1988), которые наблюдались и у другого вида рода Canis: шакала Canis aureus. Сходные случаи отмечались в Северной Америке (Van Ballenberghe, 1983). Возможно, немногочисленность такого рода данных объясняется тем, что в большинстве случаев размножающихся самок a priori идентифицируют как членов разных стай, не принимая во внимание социальную и пространственную организацию стаи. Примером этому служит работа Месье (Messier, 1985a,b), в которой приводятся данные по динамике расположения во времени границ семейного участка группы. В обоих случаях вновь образованные пары располагались внутри группы, к которой, согласно автору, они принадлежали до этого. Конечно, это утверждение небесспорно, так как речь может идти о настоящем расщеплении стаи, однако обычно такое расщепление предполагает резкое изменение конфигурации обеих оставшихся групп в целом (Jordan et al., 1967; Grande del Brío, 1991), что в данном случае не наблюдалось.
Размножение более чем одной пары с последующей постепенной ассимиляцией существующих пространственных структур без резких изменений было названо эффектом репликации, о котором Р. Гранде дель Брио (Grande del Brio, 1988, 1991) пишет: «Они (размножающиеся недоминантные пары) обычно, во всяком случае, в северо-западных областях Пиренейского полуострова, создают свой собственный очаг внутри пространственной оболочки, таким образом, происходит то, что мы называем эффектом репликации, который состоит в создании территориальных субъединиц внутри единиц высшего порядка. В центральных областях различные группы волков поочередно сменяют друг друга, и это явление носит поэтапный характер. Так, указанные центры не оказываются занятыми одними и теми же волками на неопределенное время, но, как мы убедились, бывают заняты последовательно разными группами волков, которые замещают предыдущих, когда те стареют или умирают. Это явление можно сравнить с тем, что происходит в воронке, наполненной кофейными зернами: если убрать зерна, находящиеся на самом дне, их место сразу же оказывается занято наиболее близко расположенными к ним зернами».
Это явление вписывается в сложную циклическую структуру «передачи информации», которая выходит за рамки традиционных представлений. Собственно, объяснение эффекта репликации нужно искать в матриархальной природе общества волков. На первый взгляд может показаться, что вышеописанный эффект противоречит ранее описанным моделям поведения доминантных самок, которые играют определяющую роль в структурировании очага. Если исходить из этого, предположения Р. Гранде дель Врио не выдерживают никакой критики. Однако именно матриархальный характер общества волков делает возможным возникновение эффекта репликации, - явления, которое мы подробно проанализируем ниже.
Как правило, волчица сохраняет способность к размножению до восьмидесяти лет. В момент, когда были отмечены первые фазы репликации, самки доминантных пар находились как раз приблизительно в этом возрасте. Таким образом, пространственная организация sensu lato подвергается минимальным изменениям. Отсюда вытекает внутренняя консервативная функция традиций, выработанных в каждой конкретной стае. Взрослая доминантная самка не позволит другим самкам размножаться, пока она сама сохраняет определенную репродуктивную способность. Но по мере того, как доминантная пара стареет, параллельно начинают играть ведущую роль два скрытых процесса, с одной стороны, ослабление динамической агрессивности, с другой стороны, увеличение эстральной ахронии между доминантной самкой и молодыми самками, чей эструс задерживается (Данилов и др., 1985), и присутствие волков или, реже, волчиц- помощников. Все это делает возможным возникновение начальной фазы эффекта репликации, которая предваряет смену размножающейся пары в эко-этологическом центре, гарантирующую социальный переход с возможно меньшими потерями в системе пространственно-временных отношений и, соответственно, в биологическом сигнальном поле. Появление в стае двух выводков может иметь два пути развития. Первый состоит в полигинии, редко встречающейся у волков (Harrington et al., 1982; Рябов, 1993), и обычно сопровождается агоническим поведением самок. Второй тип множественности годовых выводков описан. Поярковым и Покровским (1988) у вида Canis aureus, он характеризуется отсутствием агонического поведения между размножающимися самками в отношении связывающих их социальных (родственных) отношений.
Еще одна причина редкости явления эффекта репликации состоит в самих социальных требованиях этого понятия. Дело в том, что в волчьих популяциях, оказавшихся под давлением со стороны человека, часто наблюдается физическое устранение одного из членов доминантной пары. Это приводит к образованию разновозрастных размножающих пар (самец значительно старше самки или наоборот). При таких обстоятельствах не представляется возможным синхронное достижение обоими членами пары предела размножающейся способности. Это значит, что социальную роль доминантного самца при его смерти наследует субдоминантный самец, и при этом не возникает и не может возникнуть эффект репликации, так как в нем нет необходимости.
Итак, на основе вышеуказанных наблюдений в эффекте репликации можно выделить три следующих этапа:
1. Начальная стадия, которая характеризуется дупликацией центров размножения (один из них, старой самки, располагается в очаге, другой, молодой самки - в жизненном пространстве или в пространственной оболочке).
2. Стадия поддержания - переходная стадия, при которой продолжается размножение обеих самок, но молодая самка начинает играть все более активную роль в жизни стаи.
3. Стадия преемственности характеризуется тем, что старая самка перестает участвовать в размножении и функция репродукции полностью переходит к молодой самке. При этом основная структура пространственной организации семей не испытывает существенных изменений.
В данном случае мы приводим идеальную схему эффекта репликации, тогда как в реальных ситуациях те или иные стадии могут модифицироваться или редуцироваться.
Биологический смысл эффекта репликации заключается в безболезненном наследовании от поколения к поколению всей структуры семейного участка и
Обсуждение _________________________________________________________________________________ 195
биологического сигнального поля. На наш взгляд, принципиальной важностью этого способа перехода является сохранность адаптивных традиции данной стаи. Возможность многолетней передачи опыта через ряд поколений в наиболее полной форме осуществляется именно благодаря эффекту репликации.
Как отмечалось выше, эффект репликации наблюдается у волков довольно редко. Можно поставить вопрос о том, действительно ли данный феномен редок в природе или сама его регистрация очень затруднена для науки. Кропотливые долгосрочные исследования, требующиеся для выявления эффекта репликации, проводились в весьма ограниченном количестве, поэтому, его вероятность, скорее всего, выше, чем это отмечено в литературе. Однако следует имеет в виду, что сочетание условий, приводящее к эффекту репликации, само по себе встречается регулярно, но не часто. Еще раз подчеркнем, что антропогенное воздействие уничтожает возможность возникновения данного биологически оправданного феномена.
Выводы
Выводы
1. В семейном участке минимальных популяционных группировок волка, Canis lupus lupus L., 1758, исходя из характера и динамики пространственного распределения их особей в течение жизненного цикла, справедливо выделить три пространственные субъединицы: очаг, жизненное пространство и пространственная оболочка.
2. В процентном соотношении площадь очага у семейных участков групп волков, находящихся под влиянием разных социальных, экологических и антропогенных факторов сходна, тогда как площадь других пространственных субъединиц сходна только у семейных участков групп волков, находящихся под влиянием сходных факторов.
3. На стыках между пространственными субъединицами и по обеим сторонам буферной зоны наблюдается сгущение элементов биологического сигнального поля, градиент которого зависит от периода жизненного цикла вида.
4. Семейные группы волка можно подразделить на развито дифференцированные и несформированные, в зависимости от степени образования пространственных структур их семейных участков, от характера распределения элементов биологического сигнального поля и от характера внутригрупповых социальных связей.
| 196 |
5. В течение всего жизненного цикла продолжаются внутригрупповые взаимодействия между особями семейной группы, что свидетельствует о существовании стаи круглый год. Это особенно верно для тех групп, у которых существуют взрослые, не участвующие в размножении самцы, которых можно рассматривать как звенья внутригрупповых социальных отношений, особенно важные в весенний период.
Благодарность
Автор выражает самую искреннюю признательность своим учителям и научным руководителям Андрею Дмитриевичу Пояркову и профессору Борису Дмитриевичу Васильеву за неоценимую помощь на всех этапах создания настоящей работы, за ценные указания, предоставляющие возможность должным образом оценить свои достоинства и недостатки и постоянно учиться на своих ошибках.
В какой то мере, достижение настоящей работы также является плодом творческих усилий Рамона Гранде дель Брио, одного из наиболее одаренных специалистов по волку в Европе и в Испании в частности. За то, что он сумел зародить в авторе интерес к крупным хищникам и к проблематике, связанной с ними, и поставил на ноги будущего исследователя, заражая его энтузиазмом и научной ненасытностью, мы хотели бы выразить ему глубокую благодарность.
Полевые работы всегда связанны с множеством трудностей и препятствий, большинство из которых невозможно предугадать. Конструктивный творческий подход, во многом, определяет успех проводимых работ. В связи с этим заслуживает особого упоминания Елена Михайловна Литвинова, принявшая участие в значительной части исследований не как помощник, а как соавтор, вкладывавший в работу свои собственные усилия. Только благодаря ее тонкой интуиции, объективной строгой критике, всепогодной готовности к работе и умению создать психологический комфорт автору удалось выполнить трудоемкие задачи, которые перед ним стояли. Я хочу также выразить ей отдельную благодарность за неоценимую помощь в редактировании рукописи.
За существенную организационно-техническую помощь и глубокую заинтересованность в успехе данного исследования я хочу поблагодарить бывшего заместителя директора по науке Воронежского Биосферного Государственного заповедника, занимающего в настоящие время эту же должность в Национальном парке «Орловское полесье», В.Д. Казьмина.
За доброе отношение и неоценимую всестороннюю помощь хочу выразить свою благодарность директору государственного заповедника «Калужские засеки» Сергею Васильевичу Федосееву и его заместителю по науке Ольге Геннадьевне Червяковой, а также за энтузиазм, с которым они, вместе с коллективом сотрудников заповедника, восприняли наши исследования на территории «Калужских засек».
Автор весьма признателен сотруднику ИПЭЭ РАН В.И. Крутовой за внедрение нового и перспективного метода кинологической идентификации в нашу работу и азарт в развитии изучения волка, а также руководителю группы по этологии и поведенческой экологии наземных позвоночных ИПЭЭ РАН В.В. Рожнову за поддержку и любезное предоставление дополнительных приемников спутниковой навигационной системы (GPS).
Мне особенно приятно благодарить своего друга и коллегу C.B. Огурцова за моральную поддержку, участие и организационную помощь в проведении некоторых этапов полевых работ. Его отзывчивость и постоянную готовность к обсуждению любого вопроса трудно переоценить. Автор ему благодарен за множество плодотворных бесед на самые разные темы, в том числе, о выборе адекватного статистического аппарата.
Кроме того, хотелось бы особо поблагодарить А.Г. Николаева и других териологов научного отдела Воронежского заповедника, любезно поделившихся с нами своими чрезвычайно ценными наблюдениями, А.Б. Костина, предоставившего нам ценную информацию о фауне заповедника «Калужские засеки» и полезный картографический материал, сотрудников охраны заповедника «Калужские засеки» за понимание и содействие, главного лесничего Михайловского лесничества Национального парка «Орловское полесье» B.C. Тихонова за человеческий прием и помощь в более глубокое понимание особенности национального парка.
Многочисленные коллеги оказывали нам свою поддержку в ходе исследований, существенно помогали своими конструктивными вопросами, комментариями и тяжелым физическим трудом на различных этапах сбора полевого материала. Таким образом к нам присоединились многие аспиранты и студенты кафедры зоологии позвоночных биологического факультета МГУ, без помощи которых наша работа не получилась бы такой полной. Мы особенно признательны С.Н. Сергееву, а также считаем необходимым поблагодарить всех
Благодарность_______________________________________________________________________________ 199
студентов и аспирантов, принимавших участие в наших полевых работах: Квартального П., Кузнецова В., Кузнецову М., Фесенко Г., Ларсена Э.М., Брагину Е., Чикурову Е., Баскину С., Ястребова С., Лыкова Е., Бабицкого А., Казанского Ф., Попова В., Звычайную Е., Зарубину Т., Карагичеву Ю., Крылович О., Антоневич А., Чулкину М., Мельник К., Гатилова А., Шахпаронова В., Пояркова Н., Вострецову Е., Рогачеву А.
Отдельную сердечную благодарность мы выражаем Н.М. Пронину, егерю Воронежского заповедника, благодаря искренним заботам которого Крутовской кордон приобрел для нас особое значение.
Библиография
1. Бадридзе Я.К., 1979. Запасание пищи волками // Экологические основы охраны и рационального использование хищных млекопитающих. АН СССР. Изд-во Наука, Москва.