
5. Физико-химические св-ва белков








№1 Белки – высокомолекулярные азотсодержащие органические вещества, состоящие из аминокислот, соединённых в цепи с помощью пептидных связей, и имеющие сложную структурную организацию. Аминокислоты- мономеры белков, органические карбоновые кислоты, у которых как минимум один из атомов водорода углеводородной цепи замещён на аминогруппу. Они бывают: протеиногенные (входящие в состав белков) и не протеиногенные. Все аминокислоты белков относятся к L-аминокислотам(рецемизация), содержащим аминогруппу NH2 в альфа положении. Радикал в простейшем случае представлен атомом водорода (глицин), а может иметь и сложное строение. Поэтому альфа аминокислоты отличаются друг от друга прежде всего строением радикала, а следовательно, и физико-химическими свойствами, присущими этим радикалам. Бывают: структурная классификация (по строению радикала), электрохимическая (по кислотно-основным свойствам аминокислот), биологическая (по степени незаменимости аминокислот). Первичная структура (линейная последовательность аминокислот в пептидной цепи) закодирована в молекуле ДНК и реализуется в ходе транскрипции и трансляции. Все белки имеют уникальную для данного белка структуру. Последовательность аминокислотных остатков в пептидной цепи – форма записи некоторой информации, которая диктует пространственную укладку длинной линейной цепи в более компактную структуру или конформацию. В основе полипептидной цепи чередуются жесткие структуры (плоские пептидные группы) с относительно подвижными участками (-CHR), которые способны вращаться вокруг связей. Такие особенности строения влияют на укладку в пространстве. Вазопрессин и окситоцин 7 из 9 ам.к-ых ост идентичны, но они выполняют совершенно разную роль.Пептидная связь CO-NH
№2 Вторичная структура образуется за счёт взаим м функц группами в пепт оставе, может быть 2 типов, альфа спирали - протекает через каждые 4 аминокислоты, в одном витке 3,6 ост, бета структура - формируется между линейными областями либо одной цепью, либо двумя. Цепи бывают параллельные (NN), и антипараллельные NC.
Могут быть области с нерегулярной вторичной стр-ой, т.е беспорядочные клубки.
Третичная стр-ра - трехмерная стр-ра, образующаяся между взаимодействием разных R.
Взаимодействие - гидрофобные, ионные, водородные (слабые); дисульфидные мостики, по типу сложного эфира, простого эфира (сильные). Некоторый специфич. порядок чередов. 2 струк. наблюд. разных по стр-ре белках и называется супервторичная.
По наличию альфа спиралей и бета структур глобулярные белки можно разделить на категории: 1) имеющие только альфа спирали (гемоглобин, миоглобин); 2) имеющ. альфа спирали и бета структ, при этом альфа и бета структ. часто образуют однотипные сочетания встреч. в разных индивид. белках. (триоза фосфат изомераза) -имеет супер 2-ую структ. типа бета бочонка.; 3) имеют только вторич бета структ.(иммуноглобулин); 4) белки имеющие лишь незначит кол-во регулярных вторич. стр-р (метанмопротеины)
Характер простр. укладки пептид цепи определ аминокислотным сос-ом и чередован. остатком аминокис цепи => конформация такая же специф характеристикаиндивид белка как и первичная стр-ра.
Основы функционир белков. Центр связыв белка с лигандом или активный центр образуется на поверхности глобулы и формируется из радикалов аминокислот остатков сближ на уровне трет стр-ры. Это центр может присоед к себе другие молекулы-лиганды. Белки проявляют высокую специфичность при взаимод. с лигандом, кот обеспеч комплиментарностью стр-ры активного центра к стр-ре лиганда.
Комплиментарность-простр и хим соответст взаимод поверхностей. В основе функционир белков лежит их специфич взаимод с легандами. Доменная стр-ра и ее роль в функцион белков. Длинные полипиптид цепи складыв в несколько компактных относит независимо областей. Они имеют самост третичную стр-ру и назв доменами. Благодаря доменной стр бел легче формируется их трехмерная стр. Центры связыв белка с лигандом часто распол м/д доменами. Разные домены в белке могут перемещаться относит др друга при взаим с легандами (гексокенеза). В некоторых белках домены вып самостоят ф-ии связыв с различ лигандами = многофункц белки. Легандами могут быть не только низкомол органические и неорган молеулы но и макромолекулы (ДНК,РНК, полисахар белки).
№3 Многие белки содержат в своём составе только одну полипептидную цепь, такие белки наз. Мономерами в то же время сущ белки, сост. из 2х и более полипептид. Цепей. После формирования 3х мерной структуры какждой полипептид. цепи они объединяются с помощю тех же слабых взаимодействий, кот. участвовали в обр. третич. сруктуры: гидрофобных, ионных, водородных. Колич и взаиморасположение полипептид. цепей в пространстве наз. 4тичной структурой. Отдельные полипептидные цепи в таком белке носят наз. протомеров или субъединиц. Белок, сод. В своём составе неск. протомеров наз. олигомерным. Узнавание и присоед. отдельных протомеров олигомерного белка происходит благодаря формированию на их пов. контактных участков, кот. сост. из радикалов аминок-т, собранных в данном месте в процессе обр. третич. структуры. Совокуп. этих радикалов формир уникальные пов-ти, способ с высокой специфичностью объединяться друг с другом. Специфичность связывания контактных уч-ков опред их комплементарности – пространственным хим соответствием взаимодейств пов-тей (впадины и вывступы на пов. одной молекулы должны совпадать с выступами и впад на пов другой молекулы)
Гекоскиназа 2 протомера, гемоглобин 4 протомера, пируватдегидрогеназа 312 протомеров.
№4 Гидрофобные взаимодействия, а так же ионные и водородные связи относятся к чису слабых. Поддержание конформации возможно благодаря возникновению множ слабых связей между участками полипептид цепи. Белки сост из огромного числа атомов, наход в постоянном движении, что приводит к небольшим перемещениям отдельных участков полипептид цепи, кот обычно не наруш общую структуру белка и его ф-ции. Т.е. белки облад конформационнойц лабильностью – склонностью к небольшим изменениям конформации за счёт разрыва одних и образ других слаб связей. Конформ белка может меняться при изменении хим и физич св-в среды, а так же при взаимодействии белка с др молекулами, что играет огромную роль в функц белка в живой клетке. Разрыв большого колич слаб связей в молекуле белка приводит к разрушению её нативной информации. Так как разрыв связей ностит случ хар-р, то молекулы одного индивид. белка преобретают в р-ре форму случайно сформир беспорядочных клубков, отличающихся друг от друга трёхмерной стр-рой. Потеря нативной инф сопровождается утратой специфич ф-и белков. Этот процесс носит наз денатурация. При денатурации белков не происходит разрыва пептид связей, т.е. первич стр-ра не нарушается. Ф-ры, выз денатурацию: -выс температура >50гр, -интенсив встряхивание р-ра, приводящее к соприкосновению белковых молекул с воздуш средой на пов раздела фаз и изменению конф этих молекул, - орг в-ва (спирт, фенол, мочивина) способны взаимодейств с функц группами белков, что приводит к их конформационным изменениям, - кисл и щёлочи (изменяя pH среды выз перераспределение связей в молекуле белка), - соли тяж металлов (Cu, Hg) образ прочные связи с важными функц группами белков, - гетергенты (мыло) – в-ва, содерж гидрофобный углеводородный радикал и гидрофильную функц группу. Шапероны – белки, способные связываться с белками, находящимся в неустойчивом состоянии. Они способны стабилизировать их конформацию. Шапероны, участывующие в защите клет. белков от денатурирующих воздействий относятся к белкам светового шока(БТШ).При действии различных стрессовых факторов в клетках усиливается синтез БТШ. Имея высокое сродство к гидрофобным участкам частично денатурированных белков они могут препятствовать их полной денатурации ивосстанавливать нативную инфу белков.
5. Физико-химические св-ва белков
1 Наиболее характерными физико-химическими свойствами белков являются: высокая вязкость растворов,
незначительная диффузия, способность к набуханию в больших пределах, оптическая активность, подвижность в электрическом поле, низкое осмотическое давление и высокое онкотическое давление, способность к поглощению Уф-лучей при 280 нм (это последнее свойство, обусловленное наличием в белках ароматических аминокислот, используется для количественного определения белков). М колеблется от 6 тыс. до 1 млн. зависит от кол-ва аминокислотных остатков полипептидной цепи.
Белки, как и аминокислоты, амфотерны благодаря наличию свободных NH2-и СООН-групп и характеризуются соответственно всеми св-вами кислот и оснований.
Белки обладают явно выраженными гидрофильными свойствами. Их растворы обладают очень низким осмотическим давлением, высокой вязкостью и незначительной способностью к диффузии. Белки способны к набуханию в очень больших пределах.
С коллоидным состоянием белков связан рад характерных свойств, в частности явление светорассеяния, лежащее в основе количественного определения белков методом нефелометрии. Этот эффект используется, кроме того, в современных методах, микроскопии биологических объектов. Молекулы белка не способны проходить через, полупроницаемые искусственные мембраны (целлофан, пергамент, коллодий), а также биомембраны растительных и животных тканей, хотя при органических поражениях, например почек, капсула почечного клубочка (Шумлянского -Боумена) становится проницаемой для альбуминов сыворотки крови, и они появляются в моче.
Простые белки построены из аминокислот и при гидролизе распадаются соответственно только на аминокислоты. Сложные белки - это двухкомпонентные белки, которые состоят из какого-либо простого белка и небелкового компонента, называемого простетической группой. При гидролизе сложных белков, помимо свободных аминокислот, освобождаются небелковая часть или продукты ее распада. Простые белки в свою очередь делятся на основании некоторых условно выбранных критериев на ряд подгрупп: протамины, гистоны, альбумины, глобулины, проламины, глютелины и др.
Классификация сложных белков основана на химической природе входящего в их состав небелкового компонента. В соответствии с этим различают: фосфопротеины (содержат фосфорную кислоту), хромопротеины (в состав их входят пигменты), нуклеопротеины (содержат нуклеиновые кислоты), гликопротеины (содержат углеводы), липопротеины (содержат липиды) и металлопротеины (содержат металлы).
№6 Ионообменная хроматография: берётся колонка с ионообменной смолой. Белки связываются со смолой за счёт ионных связей, сила которых будет зависеть от заряда белков и его плотности на поверхности молекулы. Белки можно последовательно снимать с колонки заменяя их на другие катионы или анионы, используя солевые растворы разной концентрации и pH. Гель фильтрация (метод разделения белков по молекулярной массе или метод молекулярного сита): в колонке зёрна из синтетического полисахарида – декстрана, который называется серфадекс, каждое зерно – сито, которое имеет упоры определённого диаметра. Можно разделить белки по размерам, больший белок выйдет первым. Аффинная хромотография (или посродству): колонка заполняется сефарозой, в которой химически зашито какое-то вещество, которое будет реагировать только с одним белком из смеси (с которым у нее будет сродство). К ферменту зашивают субстрат, к гормону – рецептор. Затем белок снимается с колонки подходящим буферным раствором. Метод помогает выделить белки высочайшей степени очистки. Ультрафильтрация: используется толстостенная металлическая ячейка. На дне сито с разным диаметром пор. Под давлением мелкие частицы (примеси) и жидкости уходят, а белок остаётся и концентрируется, обеззараживается.
№7 В организме чела 50тыс. белков,отличающихся первичной струтрурой,конформацией,строением активного центра и функцями. Кроме того б различаются количеством амино-кт, из кот они построены. К глобулярным относ белки форма кот – эллипс. Они имеют компактную стр-ру и многие из них за счёт удаления гидрофобных радикалов внутрь хорошо р-римы в воде (глобин). Фибоиллярные белки имеют вытянутую нитевидную структуру. К ним относятся каллогены, эластин, керотин, вып в орг чел стр-рную ф-ю. Прстые белки содерж в своём составе только полипептидные цепи, сост из аминок-тных остатков. Сложные белки содерж помими полипептид цепей небелковую часть, присоед к белкут слабыми или ковалентными связями. Небелковая часть может быть представлена ионами Ме, к-либо орг молекулами и носит наз простетической группы. По сходству вып ф-ий белки можно раздеоить на группы: - ферменты (специализир белки, ускоряющие теч хим р-ий), - регуляторные белки (относятся к белковым гормонам, участвующие в поддержании гомеостаза), - рецепторные белки (сигнальные молекулы действуют на внутриклет процессы через взаимодейств с белками- рецепторами), - транспортные б (переносят специфич лиганды от одного органа к другому), стр-рные б (предают форму тканям, создают опору, опред мех св-ва), - защит б (узнают и связывают чужеродные молекулы, нейтрализуя их), - сократительные белки (при вып своих фи-й наделяют Кл способностью сокращатся и передвигаться). Белки, имеющие гомологичные участки полипептид цепи, сходную конформацию и родственные ф-и выд в сем-ва белков. Сериновые протеазы – семейство ф-тов, кот исп уникальный активированный остаток сериа, располож в актив центре, для связ и каталитич гидролиза апептид связей в б. Иммуноглобулины – специфические белки, вырабатываемые б-лимфоцитами в ответ на попадание.
№8 Биуретовый метод: белки в щелочной среде в присутствии солей меди дают фиолетовое окрашивание, интенсивность которого определяется на ФЭКе, и по калибровочной кривой определяется содержание белка. Если количество белка в сыворотке меньше 65 г/л – гипопротеинемия, если больше 85, то – гиперпротеинемия. Распространённым методом разделения белков сыворотки крови является электрофорез на бумаге, по средством которого можно разделить белки на 5 фракций: альбумины, альфа1-, альфа2-, бета и гамма глобулины. Сущность метода: электрофорезом называется процесс передвижения заряженных частиц под действием электрического поля. Разделение белков основано на различной скорости перемещения белков в электрическом поле в зависимости от знака и величины электрического заряда и молекулярной массы белка. В плазме крови содержится 7% всех белков организма, концентрация 60-80 г/л. Функции: поддержание осмотического давления (белки связывают воду и удерживают её в кровеносном русле), образуют белковую буферную систему крови и поддерживают pH в пределах 7,37-7,43; выполнят транспортную функцию, определяют вязкость крови, являются резервом аминокислот в организме, осуществляют защитную функцию. При ряде заболеваний происходит изменение соотношения белковых фракций при электрофорезе по сравнению с нормой. Такие изменения называются диспротеинемия. Содержание некоторых белков резко увеличивается при патологических состояниях, такие белки называются – белки острой фазы. К ним относятся: С-реактивный белок, альфа1 антитрипсин, гаптоглобин, кислый гликопротеин, фибриноген.
№9 Иммуноглобулины – специфические белки, вырабатываемые б-лимфоцитами в ответ на попадание в орг чужеродных стр-р, наз антигенами. Молекула сост из 4х полипептидных цепей, 2х идентичных лёгких-L, содерж ок 220 аминок-тных остатков и 2х тяжёлых-H, сост из 440 аминок-т каждая. Все 4 цепи соединены друг с другом множеством нековалентных и 4-мя дисульфидными связями. Связывание антигена с антителом участвует не вся аминок-ная последовательность обеих цепей, а всего лишь 20-30 аминок-т, расположенных в гипервариабельных облостях каждой цепи. Именно эти области определяют уникальные способности каждого клона антител взаимодействовать с соответствующим комплиментарным антигеном. Обнаружение и связ чужеродных антигенов происходит с помощью специфических антигенсвязывающих участков. Специфичность пути разрушения комплекса антиге – антитело зависи от класса антител. Существует 5 классов иммуноглобулинов: A, D, G, E, M. М – первый класс антител, развив в б-лимфацитах. Взаимодейств антигена с М изменяет его конформацию, наруш целостность мембраны и убивает клетку. G сост 75%, способен проникать через плацентарный барьер и обеспечивать внутриутробную защиту плода от инф. А – присутствует в секретах желёз орг: слюна, пищ сок. Образующийся при взаим с антигеном комплекс препятствует прикреплению антигенов к пов эпителиальных клеток и проникновению их в орг. Е – содерж в крови крайне мало. Способствует секреции БАВ (серотонин, гистамин. D – в очень малых колич.
№10 Ферменты или энзимы – белковые катализаторы, ускоряющие реакции в клетке.
Около 10000. Ферменты имеют белковую структуру, одно обнаружена способность некоторых молекул РНК осуществлять аутокатализ. Такие РНК называются рибозимы. Ферменты катализируют превращение веществ, которые называются субстратами в продукты. Ферменты отличаются от небиологических катализаторов следующими свойствами: 1.Высокой эффективностью действия, скорость ферментативной реакции в 109 – 1012 раз выше. 2.Высокой специфичностью действия, способность выбирать определенное S и катализировать специфическую реакцию. Благодаря действию ферментов реакции в клетке не беспорядочны, не перепутываются, а образуют строго определенные метаболические пути. 3.Мягкими условиями протекания ферментативной реакции. t=37, нормальное атмосферное давление, pH близкое к нейтральному. 4. Способностью к регуляции. Каталитическая активность многих ферментов может изменяться в зависимости от концентрации веществ регуляторов, что делает фермент – организатором обменных процессов в клетке.
Классификация и номенклатура. Изоферменты. Каждый фермент: рабочее (короткое для повседневного использования), систематическое (для однозначной идентификации фермента. В рабочем названии суффикс аза). Каждый класс (всего 6) состоит из многочисленных подклассов с учетом преобразуемой хим. группы субстрата, донора и акцептора преобразуемых группировок, наличия дополнительных молекул. Каждый из 6 классов имеет свой порядковый номер, закрепленный за ним: 1.Оксидоредуктазы – ферменты, катализирующие окислит.-востановит. реакции. Подразделяются на 17 подклассов. 2.Трансферазы – ферменты, катализирующие реакции переноса различных групп от одного субстрата к другому. 8 подклассов. 3.Гидролазы – ферменты, катализирующие разрыв связей в субстратах с присоединением воды. 11 подклассов. 4. Лиазы – ферменты, катализирующие реакции разрыва связей в субстрате без присоединения воды или окисления . 4 подкласса. 5.Изомеразы – ферменты, катализирующие превращения в пределах одной молекулы. 6.Лигазы – ферменты, катализирующие соединение двух молекул с использованием энергии фосфатной связи. 5 подклассов.
Изоферменты – формы фермента, который катализирует одну и туже реакцию, но различны по некоторым свойствам (аминокисл. составу, М, Составу субъединиц, электрофоретической подвижности и др.). Изоферменты – продукты экспрессии разных генов. Существуют различия в распределении изоферментов в разных тканях, в разных участках клетки, что отражает различие в метаболизме. Одним из осн-х механизмов образования изоферментов включает объединение разных субъединиц в разных комбинациях при образовании активного олигомерного фермента, т.е. 4-ная структура.
11)Зависимость скорости ферментативной реакции .
При повышении температуры на каждые 10 °С скорость увеличивается примерно вдвое (правило Вант-Гоф-фа). Однако для ферментативных реакций это правило справедливо лишь в области низких температур — до 50-60 °С. При более высоких температурах ускоряется денатурация фермента, что означает уменьшение его количества; соответственно снижается и скорость реакции. При 80-90 °С большинство ферментов денатурируется практически мгновенно. Количественное определение ферментов рекомендуется проводить при 25 °С.
Изменение рН приводит к изменению степени ионизации ионогенных групп в активном центре, а это влияет на сродство субстрата к активному центру и на каталитический механизм. Кроме того, изменение ионизации белка (не только в области активного центра) вызывает конформационные изменения молекулы фермента. существует некоторое оптимальное состояние ионизации фермента, обеспечивающее наилучшее соединение с субстратом и катализ реакции. Оптимум рН для большинства ферментов лежит в пределах от 6 до 8. Однако есть и исключения: например, пепсин наиболее активен при рН 2. Количественное определение ферментов проводят при оптимальном для данного фермента рН.
По мере увеличения времени инкубации скорость реакции снижается. Это может происходить вследствие уменьшения концентрации субстрата, увеличения скорости обратной реакции (в результате накопления продукта прямой реакции), ингибирования фермента продуктом реакции, денатурации фермента.
При увеличении количества субстрата начальная скорость возрастает. Когда фермент становится полностью насыщенным субстратом, т.е. происходит максимально возможное при данной концентрации фермента формирование фермент-субстратного комплекса, наблюдают наибольшую скорость образования продукта. Дальнейшее повышение концентрации субстрата не приводит к увеличению образования продукта, т.е. скорость реакции не возрастает.
№12 Обратимые ингибиторы связ с ферментом слабыми нековолентными связями и при опред усл легко отделяются от ф-тов. Бывают: конкурентными (обратимое снижение скорости ферментативной реккции,вызванное ингибитором,связываясь с активным центром и препятствывая образованию фермент-субстратного комплекса); неконкурентные (ингибитор взаимодействует с фер-том в участке, отличном от активного центра). Необратимое ингибирование наблюдается в случае образования ковалентных связей между молекулами ингибитора и ф-та. Модификации подвергается актив центр, и ф-т не вып кат ф-ий (ионы Ме: Hg, Ag). Многие лек препараты оказывают терапевтическое действие по мех конкурентного ингибирования. Четвертичные аммониевые основания ингибируют ацетилхолинэстеразу, катализирующюю р-ю гидролиза ацетилхолина на холин и уксусную к-ту. При добавлении ингибиторов, активность ацетилхолистеразы уменьшается, конц субстрата увелич, что сопровождается усилением проведения нервного импульса. Ингибитора холинэстеразы используются про лечении мышечных дистрофий.
№13 Большинство ф-тов имеет внутриклет локализацию и распределены в орг нереавномерно. Все ф-ты одного метаболитического пути находятся в одном отделе клетки. Особенно разделение метаболических путей важно для противоположно направленных катаболических и анаболических процессов (синтез жирных к-т происх в цитопл, а их распад в митохондриях). Энзимодиагностика. В норм условиях активность ф-тов в сыв крови относит невелика. При поражении ряда орг и тканей происх увеличение актив ф-тов. Количественно определяя нефункциональные ф-ты плазмы, можно диагностировать заболевания (при заболиваниях костей пов актив щелочной фосфатазы, при заболеваниях печени – аланинаминотрансферразы). Энзимотерапия. Исп ф-тов с терапевтической целью применяется давно. Ещё в прошлом веке, после открытия пепсина, его стали применять при лечении диспепсий и труднозаживающих язв. Более 25% всех ф-тов для проявления полной каталитической активности нуждаются в ионах металла. Ионы Ме стабилизируют молекулы субстрата. Для нек ф-тов субстратом служит комплекс превращаемого в-ва с ионом Ме (для большинства кеназ в кач одного из субстратов выст комплекс Mg-АТФ). Ионы Ме стабилизир актив центры фермента, служит мостиком между ф-том и субстратом (Mg, Mn, Zn). В отсутствии Ме нек ф-ты активностью не обладают – метпаллоэнзими (пируваткиназа). Ионы Ме выст в роли регуляторных молекул (Ca служит активатором ф-та протеинкиназы-С).
№14 Для проявления каталитической активности большинству ферментов необходимо наличие кофермента. Кофермент может быть связан с белковой частью молекулы ковалентными (липоевая кислота) и нековалентными связями (например тиаминдифосфат). Традиционно к коферментам относят производные витаминов, геммы входящие в состав цитохромов. Дегидрогеназы катализируют реакции отщепления водорода (пример: НАД и ФАД). Все ферменты этой группы обладают высокой субстратной специфичностью. Витамин В2. в основе его структуры лежит структура изоаллоксазина, соединенного со спиртом ребитолом. В слизистой оболочке кишечника после всасывания витамина происходит образование кофермента ФАД. Он входит в состав флавиновых ферментов, принимающих участие в окислит.-востанов. реакциях. Вит В6. Все его формы используются в организме для синтеза коферментов: перидоксальфосфата и передоксаминфосфата. коферменты образуются путем фосфорилирования по гидроксиметильной группе в 5-ом положении перемидинового кольца. Вит РР. Никотиновая кислота входит в организме в состав НАД, выполняющего функцию кофермента различных дегидрогеназ.
№15 Обмен веществ складывается из 3 этапов: поступление веществ в организм, межуточный обмен (тканевое превращение веществ), образование и выведение конечных продуктов. Пища человека содержит множество химических соединений, как органических, так и минеральных, они делятся на основные пищевые вещества (белки, жиры, углеводы) и минорные (витамины и минеральные соединения). Основные пищевые вещества – полимеры в ЖКТ гидрализуются при участии ферментов до мономеров, которые проникают через клеточные мембраны кишечного эпителия. Полимеры практически не всасываются. С кровью мономеры транспортируются во все органы и ткани и используются клетками. Пищевые вещества могут быть заменимыми и не заменимыми. К не заменимым относятся все минеральные соединения, витамины, некоторые аминокислоты (Валин, лейцин, изолейцин, треонин, метионин, аргенин, лизин, фенилаланин, триптофан, гистидин) и полиненасыщенные жирные кислоты (ленолевая, линоленовая). Метаболизм: выделяют 2 направления превращения веществ: катаболизм и анаболизм. При катаболизме органические вещества распадаются до СО2 и Н2О, процесс эгзэрганический (выделение энергии). У взрослого человека за сутки высвобождается 8-12 тысяч кДж. Анаболизм – превращение простых веществ в более сложные. Многие реакции анаболизма являются эндэргоническими (поглощение энергии) источником которой служит процесс катаболизма.
№16 Белки: общая суточная потребность 80-100гр из них половина должна быть животного происхождения. Любые пищевые белки сравниваются по составу аминокислот с эталоном (в качестве эталона – белок куриного яйца, наиболее отвечающий физиологическим потребностям организма). Углеводы: биологическую ценность имеют полисахариды (крахмал гликоген), дисахариды (сахароза, лактоза, мальтоза). Основная функция углеводов – энергетическая, но они выполняют структурные и др. функции. Суточная потребность 400-500 гр. из них 400 приходится на крахмал. Жиры: суточная потребность 80-100 гр. из них 20-25 гр. растительные. С жирами пищи поступают не заменимые для организма жирорастворимые витамины и витаминоподобные соединения. Вода относится к незаменимым компонентам пищи, хотя небольшие количества воды образуются из белков, жиров и углеводов при обмене их с тканями. Суточная потребность 1750-2200 гр.
Пищевые вещества могут быть заменимыми и не заменимыми. К не заменимым относятся все минеральные соединения, витамины, некоторые аминокислоты (Валин, лейцин, изолейцин, треонин, метионин, аргенин, лизин, фенилаланин, триптофан, гистидин) и полиненасыщенные жирные кислоты.
№17 Витамины: их концентрация и суточная потребность невелики, но при недостаточном поступлении в организм наступают характерные и опасные патологические изменения. Бывают: 1) Жирорастворимые (А, Д, Е. К) и водорастворимые (группа В). 2) Витамины выполняющие в организме коферментные функции (группа В), витамины – гормоны (А, Д, К) и витамины-антиоксиданты (С, Е). Гиповитаминозы возникают из-за: недостатка пищи, дисбактериоза и бывают наследственной формы гиповитаминоза (дефект ферментов, превращение витаминов в кофермент, отсутствие синтеза апоферментов). Гипервитаминозы – избыточное потребление приводит к нарушениям обмена и функций организма.
№18 Минеральные вещ-ва поступают в организм с пищей и водой 90% живых клеток C,H2,O2,N. Классификация: 1. Макроэлементы: Na,Cl,P,Ca,Mg,K,S – составляют 9% веса тела; 2. Микроэлементы: Fe,I,Cu,Mn,Zn,Co,Hb,Se,Vd,Ni; 3. Ультрамикроэлементы: все остальные. Роль: I Макроэлементов: Ме этой группы используются клетки для создания электрич биопотенциалов и биотопов, а также в качестве «спусковых крючков» опосредущих передачу сигналов. II Микроэлементы: 1.входит в состав актив центров различ ферментов; 2.входят в состав прогармонов и активных гармонов; 3.входят в состав транс белков (Fe в гемоглобин); 4.входят в состав редокс систем, участвующих в продукции анактивации кислородных радикалов в организме. Недостаток или избыток поступления мин вещ-в в организм приводит к развитию патол процессов.
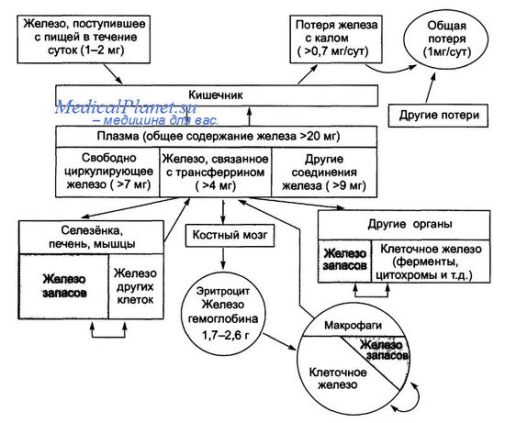
№19 Обмен железа. В ор-ме чел-ка содер-ся 3-4 гр Fe, 70%-в Нв эритроцитов, 20%-в гемоглобине мышц, до 15% в печени и селезенке, около 1% в составе гиминовых ферменов, а также белков содержит не геминовое. В процессе эволюции возникли белки способные поддерживать Fe в форме удобной для транспартировки и использовании при синтезе гемма. Это белки трансферин, феритин. Трансферин – гликопротеин плазмы крови, имеет 2 центра связывания Fe, в составе трансферина Fe3Т в форме карбонана. Трасферин содержащий Fe, эндоцитируется в клетки при участии мембранных рецепторов. Главная ф-ция перенос Fe с током крови к местам дипонирования и использования. Содержание трансферрина в плазме крови 4 гр на 1 литр. Ферритин – крупый белок, молек м=450 тыс, содержит 24 едентичных протомера, образующих полую сферу кот=12нм, в белковой оболочке 6 каналов ведущих в полость через них проникают ионы Fe, образуя железное ядро молекулы кол-во атомов Fe от 0 до 4500. Содержится в виде формы гидроксит фосфата Fe3+ . ф-ция: депонирование Fe. Его много в печени, селезенке, кост мозге. Fe освобождается из гемма при распаде эритроцитов и используется повторно.1 мг в сутки теряется с желчью, а суточное потребление 10-20 мг. Всасывание Fe в кишечнике весьма ограничено и происходит при участии белка, сходного с трансферином. Затем Fe поступает на трансфеорин крови, кот передает его феретину в клетках разных органов. В соединении с белками Fe3+, но при переходе с одного белка на другой, валентность каждый раз дважды меняется Fe3+àFe2+àFe3+ Этот процесс катализируется он-вост ферментами или самими белками переносчиками и необходимым для освобождения Fe и соединения с белком.
Общая схема обмена Fe
№20 биогеохим провинции- регионы биосферы в пределах которых по недостат или избыт определенного хим элемента выделяется естественные геохим аномалии. В Чел обл распространены болезни связанные с недостатком I (эндемический зоб). Нехватка йода приводит к базедовой болезни, угнетает деятельность центральной нервной системы, снижает эмоциональный тонус. Симптомами недостатка йода в организме являются плохое самочувствие, снижение работоспособности. У детей при недостатке йода наблюдаются отставание в росте, отклонения в умственном развитии. Ежедневная норма потребления йода – 0.1-0.2 миллиграмма. Йодом богаты, прежде всего, морепродукты – морская рыба, кальмары, креветки, мидии, морская капуста. Содержится йод и в наших повседневных продуктах: сыре, мясе, животном жире, яйцах, грибах, зерновых продуктах.
Селен является синергистом витамина Е и способствует повышению его антиоксидантной активности. Селен входит в состав фермента - йодтиронин-5-дейодиназы (контролирующего образование трийодтиронина состав белков мышечной ткани и, что особенно важно, белков миокарда. В виде селенпротеина является составной частью тестикулярной ткани. Поэтому дефицит селена приводит к ослаблению антиоксидантного статуса, антикацерогенной защиты, обусловливает миокардиодистрофию, нарушение сексуальной функции, иммунодефицита. Помимо этого селен проявляет антимутагенный, антитератогенный, радиопротекторный эффекты, стимулирует антитоксическую защиту, нормализует обмен нуклеиновых кислот и белков, улучшает репродуктивную функцию, нормализует обмен эйкозаноидов (простагландинов, простациклинов, лейкотриенов), регулирует функцию щитовидной и поджелудочной желез. В силу изложенного селен относится к геропротекторам.
21. Порфирины. Основу стр-ры простетической группы большинства гемосодержащих белков составляет порфиновое кольцо, являющееся производным порфина, котоый состоит из 4 замещенных пирролов, соединенных мужду собой метиновыми мостиками.
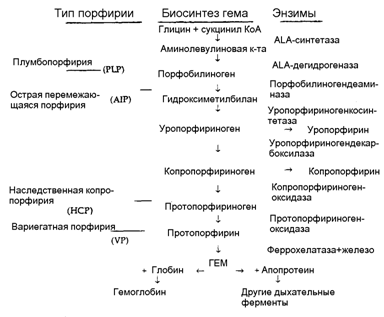
Порфирия- нарушение синтеза гемма , сопровождающееся накоплением порфириногенов и продуктов их распада в тканях, органах и крови, вызывая при этом патологию.
Наследственные- дефекты ферментов синтеза гемма(кроме АЛК-синтазы)Накопление АЛК, ПБГ, порфириногенов.
Эритропоэтические- накопление порфиринов в эритроцитах( всегда наследственные).
Болезнь гюнтера— тип наследования рецессивный.дефицит уропорфириноген-Ш-юсинтетазы, в связи с чем увеличивается неферментативный синтез уропорфириногена 1 и, соответственно, копропорфириногена. Наблюдается тяжелый фотодерматит, неэффективный эритропоэз и гемолитическая анемия, спленомегалия, накопление порфириногенов в зубах, которые приобретают розово-коричневую о фаску и светятся в темноте, моча больных светится красным светом в ультрафиолете (начиная с400нм.Заболевание проявляется в раннем детском возрасте в виде яр ко выраженной светобоязни, светящихся зубови из-за тяжелой анемии бессознательном желании употреблять больше мяса, а вдеревнях и свежей крови животных при их забое.Возможно, что в основе средневековых легенд о людях-вампирах.