
Н-Сер – Тир – Сер – Мет – Глу – Гис – Фен – Арг – Трп – Гли – Лиз – Про – Вал – Гли –








– Лиз – Лиз – Aрг – Aрг – Про – Вал – Лиз – Вал – Тир – Про – Асп – Ала – Гли – Глу –
– Асп – Глн – Сер – Ала – Глу – Ала – Фен – Про – Лей – Глу – Фен-ОН
Данные о механизме действия АКТГ на синтез стероидных гормонов свидетельствуют о существенной роли аденилатциклазной системы. Предполагают, что АКТГ вступает во взаимодействие со специфическими рецепторами на внешней поверхности клеточной мембраны (рецепторы представлены белками в комплексе с другими молекулами, в частности с сиаловой кислотой). Сигнал затем передается на фермент аденилатциклазу, расположенную на внутренней поверхности клеточной мембраны, которая катализирует распад АТФ и образование цАМФ. Последний активирует протеинкиназу, которая в свою очередь с участием АТФ осуществляет фосфорилирование холинэстеразы, превращающей эфиры холестерина в свободный холестерин, который поступает в митохондрии надпочечников, где содержатся все ферменты, катализирующие превращение холестерина в кортикостероиды. Соматотропный гормон (СТГ, гормон роста, соматотропин) синтезируется в ацидофильных клетках передней доли гипофиза; концентрация его в гипофизе составляет 5–15 мг на 1 г ткани. СТГ человека состоит из 191 аминокислоты и содержит две дисульфидные связи; N- и С-концевые аминокислоты представлены фенилаланином. СТГ обладает широким спектром биологического действия. Он влияет на все клетки организма, определяя интенсивность обмена углеводов, белков, липидов и минеральных веществ. Он усиливает биосинтез белка, ДНК, РНК и гликогена и в то же время способствует мобилизации жиров из депо и распаду высших жирных кислот и глюкозы в тканях. Помимо активации процессов ассимиляции, сопровождающихся увеличением размеров тела, ростом скелета, СТГ координирует и регулирует скорость протекания обменных процессов. Многие биологические эффекты этого гормона осуществляются через особый белковый фактор, образующийся в печени под влиянием гормона – соматомедин. По своей природе он оказался пептидом с мол. массой 8000. Тиреотропный гормон (ТТГ, тиротропин) является сложным гликопротеином и содержит, кроме того, по две α- и β-субъединицы, которые в отдельности биологической активностью не обладают: мол. масса его около 30000. Тиротропин контролирует развитие и функцию щитовидной железы и регулирует биосинтез и секрецию в кровь тиреоидных гормонов. Полностью расшифрована первичная структура α- и β-субъединиц тиротропина: α-субъединица, содержащая 96 аминокислотных остатков; β-субъединица тиротропина человека, содержащая 112 аминокислотных остатков, К гонадотропным гормонам (гонадотропины) относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий гормон (ЛГ, лютропин). Оба гормона синтезируются в передней доле гипофиза и являются сложными белками – гликопротеинами с мол. массой 25000. Они регулируют стероидо - и гаметогенез в половых железах. Фоллитропин вызывает созревание фолликулов в яичниках у самок и сперматогенез – у самцов. Лютропин у самок стимулирует секрецию эстрогенов и прогестерона, как и разрыв фолликулов с образованием желтого тела, а у самцов – секрецию тестостерона и развитие интерстициальной ткани. Биосинтез гонадотропинов, как было отмечено, регулируется гипоталамическим гормоном гонадолиберином. Лютропин состоит из двух α- и β-субъединиц: α-субъединица гормона содержит из 89 аминокислотных остатков с N-конца и отличается природой 22 аминокислот.
29. Гормоны задней доли гипофиза: вазопрессин, окситоцин. Химическая природа. Механизм их действия, биологический эффект. Нарушения функций организма, связанные с недостатком выработки этих гормонов.
Гормоны вазопрессин и окситоцин синтезируются рибосомальным путем. Оба гормона представляют собой нонапептиды следующего строения: Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в положении 3 от N-конца фенилаланин вместо изолейцина и в положении 8 – аргинин вместо лейцина. Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки при родах и мышечных волокон вокруг альвеол молочных желез, что вызывает секрецию молока. Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная роль его в организме сводится к регуляции водного обмена, откуда его второе название антидиуретического гормона. В небольших концентрациях (0,2 нг на 1 кг массы тела) вазопрессин оказывает мощное антидиуретическое действие – стимулирует обратный ток воды через мембраны почечных канальцев. В норме он контролирует осмотическое давление плазмы крови и водный баланс организма человека. При патологии, в частности атрофии задней доли гипофиза, развивается несахарный диабет – заболевание, характеризующееся выделением чрезвычайно больших количеств жидкости с мочой. При этом нарушен обратный процесс всасывания воды в канальцах почек.
Окситоцин
Вазопрессин
30. Гормоны щитовидной железы: трийодтиронин и тироксин. Химическая природа, биосинтез. Механизм действия гормонов на молекулярном уровне, биологический эффект. Изменение обмена вещества при гипертиреозе. Механизм возникновения эндемического зоба и его предупреждение.
Тироксин и трийодтиронин – основные гормоны фолликулярной части щитовидной железы. Помимо этих гормонов (биосинтез и функции которых будут рассмотрены ниже), в особых клетках – так называемых парафолликулярных клетках, или С-клетках щитовидной железы, синтезируется гормон пептидной природы, обеспечивающий постоянную концентрацию кальция в крови. Он получил название ≪кальцитонин≫. Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Из L-тиронина легко синтезируется гормон щитовидной железы тироксин, содержащий в 4 положениях кольцевой структуры йод Биологическое действие гормонов щитовидной железы распространяется на множество физиологических функций организма. В частности, гормоны регулируют скорость основного обмена, рост и дифференцировку тканей, обмен белков, углеводов и липидов, водно-электролитный обмен, деятельность ЦНС, пищеварительного тракта, гемопоэз, функцию сердечнососудистой системы, потребность в витаминах, сопротивляемость организма инфекциям и др. Гипофункция щитовидной железы в раннем детском возрасте приводит к развитию болезни, известной в литературе как кретинизм. Помимо остановки роста, специфических изменений кожи, волос, мышц, резкого снижения скорости процессов обмена, при кретинизме отмечаются глубокие нарушения психики; специфическое гормональное лечение в этом случае не дает положительных результатов. Повышенная функция щитовидной железы (гиперфункция) вызывает развитие гипертиреоза
L-тироксин L-3,5,3'-трийодтиронин
31. Гормоны коры надпочечников: глюкокортикоиды, минералокортикоиды. Химическая природа. Механизм действия на молекулярном уровне. Их роль в регуляции углеводного, минерального, липидного и белкового обмена.
В зависимости от характера биологического эффекта гормоны коркового вещества надпочечников условно делят на глюкокортикоиды (кортикостероиды, оказывающие влияние на обмен углеводов, белков, жиров и нуклеиновых кислот) и минералокортикоиды (кортикостероиды, оказывающие преимущественное влияние на обмен солей и воды). К первым относятся кортикостерон, кортизон, гидрокортизон (кортизол), 11-дезоксикортизол и 11-дегидрокортикостерон, ко вторым – дезоксикортикостерон и альдостерон. В основе их структуры, так же как и в основе строения холестерина, эргостерина, желчных кислот, витаминов группы D, половых гормонов и ряда других веществ, лежит конденсированная кольцевая система циклопентанпергидрофенантрена. Глюкокортикоиды оказывают разностороннее влияние на обмен веществ в разных тканях. В мышечной, лимфатической, соединительной и жировой тканях глюкокортикоиды, проявляя катаболическое действие, вызывают снижение проницаемости клеточных мембран и соответственно торможение поглощения глюкозы и аминокислот; в то же время в печени они оказывают противоположное действие. Конечным итогом воздействия глюкокортикоидов является развитие гипергликемии, обусловленной главным образом глюконеогенезом. Минералокортикоиды (дезоксикортикостерон и альдостерон) регулируют главным образом обмен натрия, калия, хлора и воды; они способствуют удержанию ионов натрия и хлора в организме и выведению с мочой ионов калия. По-видимому, происходит обратное всасывание ионов натрия и хлора в канальцах почек в обмен на выведение других продуктов обмена,
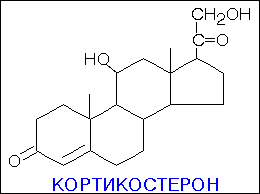
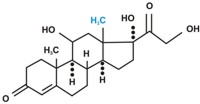
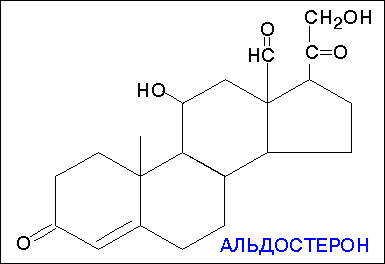
кортизол
32. Паратгормон и кальцитонин. Химическая природа. Механизм действия на молекулярном уровне. Влияние на обмен кальция, гиперкальциемия и гипокальциемия.
К гормонам белковой природы относится также паратиреоидный гормон (паратгормон). Они синтезируются паращитовидными железами. Молекула паратгормона быка содержит 84 аминокислотных остатка и состоит из одной полипептидной цепи. Выяснено, что паратгормон участвует в регуляции концентрации катионов кальция и связанных с ними анионов фосфорной кислоты в крови. Биологически активной формой считается ионизированный кальций, концентрация его колеблется в пределах 1,1–1,3 ммоль/л. Ионы кальция оказались эссенциальными факторами, не заменимыми другими катионами для ряда жизненно важных физиологических процессов: мышечное сокращение, нервно-мышечное возбуждение, свертывание крови, проницаемость клеточных мембран, активность ряда ферментов и т.д. Поэтому любые изменения этих процессов, обусловленные длительным недостатком кальция в пище или нарушением его всасывания в кишечнике, приводят к усилению синтеза паратгормона, который способствует вымыванию солей кальция (в виде цитратов и фосфатов) из костной ткани и соответственно к деструкции минеральных и органических компонентов костей. Другой орган-мишень паратгормона – это почка. Паратгормон уменьшает реабсорбцию фосфата в дистальных канальцах почки и повышает канальцевую реабсорбцию кальция. В особых клетках – так называемых парафолликулярных клетках, или С-клетках щитовидной железы, синтезируется гормон пептидной природы, обеспечивающий постоянную концентрацию кальция в крови - кальцитонин. Формула:
Кальцитонин содержит дисульфидный мостик (между 1-м и 7-м аминокислотными остатками) и характеризуется N-концевым цистеином и С-концевым пролинамидом. Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Таким образом, постоянство уровня кальция в крови человека и животных обеспечивается главным образом паратгормоном, кальцитриолом и кальцитонином, т.е. гормонами как щитовидной и паращитовидных желез, так и гормоном – производным витамина D3. Это следует учитывать при хирургических лечебных манипуляциях на данных железах.
33. Гормоны мозгового слоя надпочечников – катехоламины: адреналин и норадреналин. Химическая природа и биосинтез. Механизм действия гормонов на молекулярном уровне, их роль в регуляции обмена углеводов, жиров и аминокислот. Нарушения обмена при заболеваниях надпочечников.
Эти гормоны по строению напоминают аминокислоту тирозин, от которого они отличаются наличием дополнительных ОН-групп в кольце и у β-углеродного атома боковой цепи и отсутствием карбоксильной группы.
Адреналин Норадреналин Изопропиладреналин
В мозговом веществе надпочечников человека массой 10 г содержится около 5 мг адреналина и 0,5 мг норадреналина. Содержание их в крови составляет соответственно 1,9 и 5,2 нмоль/л. В плазме крови оба гормона присутствуют как в свободном, так и в связанном, в частности, с альбуминами состоянии. Небольшие количества обоих гормонов откладываются в виде соли с АТФ в нервных окончаниях, освобождаясь в ответ на ихраздражение. Кроме того, все они оказывают мощное сосудосуживающее действие, вызывая повышение артериального давления, и в этом отношении действие их сходно с действием симпатической нервной системы. Известно мощное регулирующее влияние этихгормонов на обмен углеводов в организме. Так, в частности, адреналин вызывает резкое повышение уровня глюкозы в крови, что обусловлено ускорением распада гликогена в печени под действием фермента фосфорилазы. Гипергликемическйй эффект норадреналина значительно ниже – примерно 5% от действия адреналина. Параллельно отмечаются накопление гексозофосфатов в тканях, в частности в мышцах, уменьшение концентрации неорганического фосфата и повышение уровня ненасыщенных жирных кислот в плазме крови. Имеются данные о торможении окисления глюкозы в тканях под влиянием адреналина. Это действие некоторые авторы связывают с уменьшением скорости проникновения (транспорта) глюкозы внутрь клетки. Известно, что и адреналин, и норадреналин быстро разрушаются в организме; с мочой выделяются неактивные продукты их обмена, главным образом в виде 3-метокси-4-оксиминдальной кислоты, оксоадренохрома, метоксинорадреналина и метоксиадреналина. Эти метаболиты содержатся в моче преимущественно в связанной с глюкуроновой кислотой форме. Ферменты, катализирующие указанные превращения катехоламинов, выделены из многих тканей и достаточно хорошо изучены, в частности моноаминоксидаза (МАО), определяющая скорость биосинтеза и распада катехоламинов, и катехолметилтрансфераза, катализирующая главный путь превращения адреналина, т.е. о-метилирование за счет S-аденозилметионина. Приводим структуру двух конечных продуктов распада
34. Глюкагон и инсулин. Химическая природа, биосинтез инсулина. Механизм действия этих гормонов на молекулярном уровне. Их роль в регуляции обмена углеводов, жиров, аминокислот. Биохимические нарушения при сахарном диабете.
 Инсулин, получивший свое название от наименования панкреатических островков. Молекула инсулина, содержащая 51 аминокислотный остаток, состоит из двух полипептидных цепей, соединенных между собой в двух точках дисульфидными мостиками. В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови. При недостаточной секреции инсулина развивается специфическое заболевание – сахарный диабет. Физиологические эффекты инсулина: Инсулин — единственный гормон, снижающий содержание глюкозы в крови, это реализуется через:
Инсулин, получивший свое название от наименования панкреатических островков. Молекула инсулина, содержащая 51 аминокислотный остаток, состоит из двух полипептидных цепей, соединенных между собой в двух точках дисульфидными мостиками. В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови. При недостаточной секреции инсулина развивается специфическое заболевание – сахарный диабет. Физиологические эффекты инсулина: Инсулин — единственный гормон, снижающий содержание глюкозы в крови, это реализуется через:
§ усиление поглощения клетками глюкозы и других веществ;
§ активацию ключевых ферментов гликолиза;
§ увеличение интенсивности синтеза гликогена — инсулин форсирует запасание глюкозы клетками печени и мышц путём полимеризации её в гликоген;
§ уменьшение интенсивности глюконеогенеза — снижается образование в печени глюкозы из различных веществ
Анаболические эффекты
§ усиливает поглощение клетками аминокислот (особенно лейцина и валина);
§ усиливает транспорт в клетку ионов калия, а также магния и фосфата;
§ усиливает репликацию ДНК и биосинтез белка;
§ усиливает синтез жирных кислот и последующую их этерификацию — в жировой ткани и в печени инсулин способствует превращению глюкозы в триглицериды; при недостатке инсулина происходит обратное — мобилизация жиров.
Антикатаболические эффекты
§ подавляет гидролиз белков — уменьшает деградацию белков;
§ уменьшает липолиз — снижает поступление жирных кислот в кровь.
Глюкагон — гормон альфа-клеток островков Лангерганса поджелудочной железы. По химическому строению глюкагон является пептидным гормоном. Молекула глюкагона состоит из 29 аминокислот и имеет молекулярный вес 3485. Первичная структура молекулы глюкагона следующая:
NH 2 - His - Ser - Gln - Gly - Thr - Phe - Thr - Ser - Asp - Tyr - Ser - Lys - Tyr - Leu - Asp - Ser - Arg - Arg - Ala - Gln - Asp - Phe - Val - Gln - Trp - Leu - Met - Asn - Thr - COOH
Механизм действия глюкагона обусловлен его связыванием со специфическими глюкагоновыми рецепторами клеток печени. Это приводит к повышению опосредованной G-белком активности аденилатциклазы и увеличению образования цАМФ. Результатом является усиление катаболизма депонированного в печени гликогена. Глюкагон оказывает сильное инотропное и хронотропное действие на миокард вследствие увеличения образования. Результатом является повышение артериального давления, увеличение частоты и силы сердечных сокращений
35. Половые гормоны: андрогены, эстрогены. Химическая природа. Механизм действия на молекулярном уровне, влияние на обмен веществ. Нарушения обмена при недостатке и избытке этих гормонов в организме.
Женские половые гормоны. Основным местом синтеза женских половых гормонов – эстрогенов – являются яичники и желтое тело. В настоящее время открыты 2 группы женских половых гормонов, различающихся своей химической структурой и биологической функцией: эстрогены (главный представитель – эстрадиол) и прогестины (главный представитель – прогестерон). Наиболее активный эстроген – эстрадиол, синтезируется преимущественно в фолликулах; два остальных эстрогена являются производными эстрадиола и синтезируются также в надпочечниках и плаценте. Все эстрогены состоят из 18 атомов углерода. Секреция эстрогенов и прогестерона яичником носит циклический характер, зависящий от фазы полового цикла: в первой фазе цикла синтезируются в основном эстрогены, а во второй – преимущественно прогестерон. Основная биологическая роль эстрогенов и прогестерона, синтез которых начинается после наступления половой зрелости, заключается в обеспечении репродуктивной функции организма женщины. В этот период они вызывают развитие вторичных половых признаков и создают оптимальные условия, обеспечивающие возможность оплодотворения яйцеклетки после овуляции. Прогестерон выполняет в организме ряд специфических функций: подготавливает слизистую оболочку матки к успешной имплантации яйцеклетки в случае ее оплодотворения, а при наступлении беременности основная роль – сохранение беременности; оказывает тормозящее влияние на овуляцию и стимулирует развитие ткани молочной железы. Эстрогены оказывают анаболическое действие на организм, стимулируя синтез белка. Мужские половые гормоны. Биосинтез андрогенов осуществляется главным образом в семенниках и частично в яичниках и надпочечниках Биологическая роль андрогенов в мужском организме в основном связана с дифференцировкой и функционированием репродуктивной системы, причем в отличие от эстрогенов андрогенные гормоны уже в эмбриональном периоде оказывают существенное влияние на дифференцировку мужских половых желез, а также других тканей, определяя характер секреции гонадотропных гормонов у взрослых. Во взрослом организме андрогены регулируют развитие мужских вторичных половых признаков, сперматогенез в семенниках и т.д.
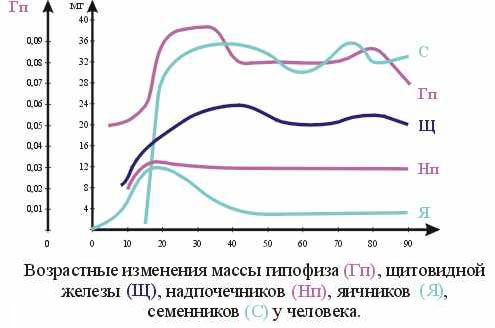
36. Неравномерность созревания и функционирования желез внутренней секреции у ребенка в различные возрастные периоды. Отличие общего гормонального фона ребенка по сравнению с организмом взрослых.
37. Мембраны, их значение для клеток и клеточных включений. Липидный бислой как основа плазматических и других клеточных мембран. Однотипичность их строения. Гликолипиды и белки мембран. Физико-химические свойства мембраны. Липидный и фосфолипидный спектр мембраны. ПОЛ – важный физиологический процесс. Изменения ПОЛ при патологии, пути защиты мембраны от ПОЛ.
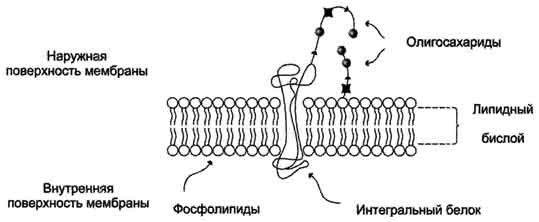
Клеточная мембрана отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды. Биологические мембраны представляют собой "ансамбли" липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий. Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы "растворены" в липидном бислое. Фосфолипиды. Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран - фосфатидилхолины и фосфатидилэтаноламины. В мембранах эукариотических клеток обнаружено огромное количество разных фосфолипидов, причём они распределены неравномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных "головок", так и ацильных остатков. Гликолипиды. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа - углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у первого углеродного атома церамида. В зависимости от длины и строения углеводной части различают цереброзиды, содержащие моно - или олигосахаридный остаток, и ганглиозиды, к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-ацетилнейраминовую кислоту (NANA). Специфические свойства биологических мембран. Благодаря указанным особенностям биологические мембраны имеют присущие им характерные черты. Они образуют протяженные бислойные структуры малой толщины (6–10 нм), объединяющие белковые и липидные компоненты с различными свойствами. Целостная структура мембраны создается за счет гидрофобных и электростатических взаимодействий, а не за счет ковалентных связей между составляющими ее молекулами белков и липидов. Гидрофобный липидный бислой представляет естественную преграду для проникновения полярных молекул. Мембраны асимметричны по своему исходному строению, что обеспечивает градиент кривизны и спонтанное образование замкнутых структур. Мембранный бислой обладает относительно малой микровязкостью. Другими словами, мембраны рыхло упакованы, что позволяет отдельным компонентам проявлять высокую подвижность в латеральном направлении. Наружные мембраны клеток отличаются от внутренних по липидному составу (последние почти не содержат стеринов, имеют соотношение ФХ/ФЭ > 1) и обладают специфическим набором ферментов и рецепторов.
Как правило, белки плазматических мембран со стороны внеклеточной среды обильно гликозилированы. Внутриклеточные мембраны содержат мало гликопротеинов и гликолипидов и характеризуются меньшей микро- вязкостью. Благодаря этому они могут образовывать органеллы малого размера. Мембранные белки выполняют различные специфические функции: рецепторные, транспортные, ферментативные, энергопреобразующие и т.д.
38. Функции мембраны: барьерная, защитная, регуляторная, энергетическая. Транспортная функция мембраны. Виды транспорта: пассивный, активный, цитоз. Ферментный спектр мембраны: транспортные АТФ-азы, механизм их действия. Повреждение мембраны – неизбежный этап любой клеточной патологии. Атеросклероз
· барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
· матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
· механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
· энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
· рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами.
ферментативная — мембранные белки нередко являются ферментами.
· Транспортная функция является одной из важных функций клеточных мембран. Мембрана создает
существенные ограничения для проникновения различных веществ, однако она не является полностью непроницаемой: небольшие нейтральные молекулы могут проникать через бислой в области структурных дефектов. Этот процесс осуществляется по градиенту концентрации переносимого вещества - из области, где его содержание высоко, в область с более низким содержанием. Такой процесс называется простой диффузие й, он осуществляется неизбирательно и с низкой скоростью. При облегченной диффузии вещества также переносятся в направлении их концентрационного градиента, но с использованием специальных структур - переносчиков или каналов, увеличивающих скорость и специфичность переноса. Известны высокоспецифические транслоказы - белковые молекулы, переносящие адениловые нуклеотиды через внутреннюю мембрану митохондрий: Nа+/Са2+-обменник - белок, входящий в состав плазматических мембран многих клеток; низкомолекулярный пептид бактериального происхождения валиномицин - специфический переносчик для ионов К+. Активный транспорт веществ осуществляется такими же механизмами, но протекает против концентрационного градиента и для своего осуществления должен быть сопряжен с энергодающим процессом. Основным источником энергии для активно- го транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы. Примером систем активного транспорта ионов является Na+/K+- АТФаза плазматических мембран животных клеток, которая ≪выкачивает≫ из клетки ионы натрия в обмен на ионы калия, затрачивая на выполнение этой работы АТФ в стехиометрии 3Nа+/2К+/1АТФ. Са2+-АТФаза осуществляет активный транспорт кальция через мембрану со стехиометрией 2Са2+/1АТФ. В так называемых сопрягающих мембранах имеются протонные насосы, работающие как Н+-АТФазы. В результате их функционирования на мембране возникает разность концентраций протонов (ΔрН) и разность электрических потенциалов. Атеросклероз — хроническое заболевание артерий эластического и мышечно-эластического типа, возникающее вследствие нарушения липидного обмена и сопровождающееся отложением холестерина и некоторых фракций липопротеидов в интиме сосудов. Отложения формируются в виде атероматозных бляшек. Последующее разрастание в них соединительной ткани (склероз), и кальциноз стенки сосуда приводят к деформации и сужению просвета вплоть до облитерации (закупорки). Атеросклероз сосудов ведет к развитию ишемической болезни сердца.
39. Слабость антиоксидантной защиты у мембран у детей ранних сроков жизни и ее последующая активация.
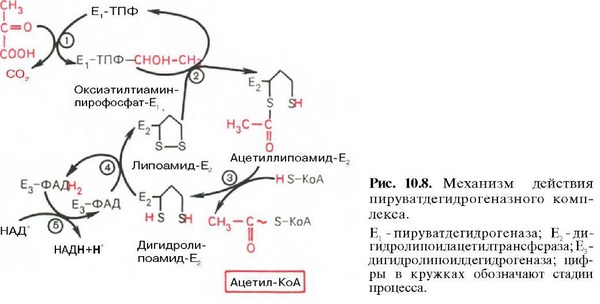
40. Окислительное декарбоксилирование пировиноградной кислоты: последовательность реакций, строение пируватдегидрогеназного комплекса. Нарушение его функции при атеросклерозе
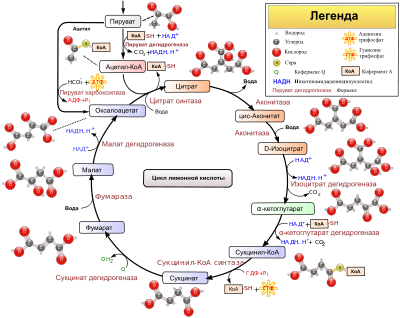
41. Цикл лимонной кислоты: последовательность реакций и характеристика ферментов. Связь между циклом трикарбоновых кислот и цепью переноса электронов и протонов. Аллостерическая регуляция цитратного цикла, его анаболические функции.
42. Понятие об обмене веществ и метаболических путях. Стадии катаболизма, взаимосвязь их с анаболическими процессами. Основные конечные продукты метаболизма у человека.
Метаболизм или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты. Обмен веществ происходит между клетками организма и межклеточной жидкостью, постоянство состава которой поддерживается кровообращением: за время прохождения крови в капиллярах через проницаемые стенки капилляров плазма крови 40 раз полностью обновляется с интерстициальной жидкостью. Серии химических реакций обмена веществ называют метаболическими путями, в них при участии ферментов одни биологически значимые молекулы последовательно превращаются в другие. Основные метаболические пути и их компоненты одинаковы для многих видов. Например, некоторые карбоновые кислоты, являющиеся интермедиатами цикла трикарбоновых кислот присутствуют во всех организмах Катаболические, анаболические пути. Ферментативное расщепление основных питательных веществ, а именно углеводов, жиров и белков, происходит в клетке через ряд последовательных ферментативных реакций. Катаболизм основных питательных веществ включает три главные стадии. На первой стадии крупные пищевые молекулы расщепляются на составляющие их основные строительные блоки. Полисахариды, например, расщепляются до гексоз или пентоз, липиды — до жирных кислот, глицерина и других компонентов, белки — до аминокислот, которых имеется 20 видов. На второй стадии большое число продуктов, образовавшихся на первой стадии, превращается в более простые молекулы, число типов которых сравнительно невелико. Так, гексозы, пентозы и глицерин, разрушаясь, превращаются сначала в трехуглеродный фосфорилированный сахар — глицеральдегид-3-фосфат, а затем расщепляются далее до единственной двууглеродной формы — ацетильной группы, входящей в состав ацетил- КоА. Двадцать различных аминокислот также дают при расщеплении лишь несколько конечных продуктов, а именно ацетил-КоА, α-кетоглутарат, сукцинат, фумарат и оксалоацетат. Продукты, образовавшиеся на второй стадии, вступают в третью стадию, которая для них всех является общей и на которой они в конечном счете окисляются до CO2 и воды. Процесс анаболизма также включает три стадии. Исходными веществами, или строительными блоками служат для него соединения, поставляемые третьей стадией катаболизма. Таким образом, третья стадия катаболизма является в то же время первой, исходной, стадией анаболизма. Синтез белка, например, начинается на этой стадии с α - кетокислот, являющихся предшественниками α -аминокислот. На второй стадии анаболизма α -кетокислоты аминируются аминогруппой доноров с образованием α -аминокислот, а на третьей, заключительной, стадии аминокислоты объединяются в пептидные цепи. Продукты метаболизма выводятся с мочой, фекалиями, выдыхаемым воздухом и потом. Специфические вещества задерживаются или удаляются из организма в той степени, в какой это необходимо для поддержания гомеостаза, при этом вместе со шлаковыми продуктами распада выводятся потенциально полезные вещества. Небольшие количества этих веществ выводятся из организма в виде кишечных газов, волос, ногтей, десквамированного эпителия кожи, кожного сала, ушной серы, слизи из полости носа и влагалища, слюны, слез, семенной жидкости и менструальных выделений. Уровни потерь этих веществ опубликованы в работах.
43. Митохондриальное окисление. Дегидрирование субстратов и окисление водорода как источник энергии для синтеза АТФ. Структура и функции переноса электронов, окислительно-восстановительный потенциал структурных компонентов цепи. Нарушение функции цепи переноса электронов при гипоксиях и других патологических состояниях.
Система митохондриального окисления - мультиферментная система, постепенно транспортирующая протоны и электроны на кислород с образованием молекулы воды. Все ферменты митохондриального окисления встроены во внутреннюю мембрану митохондрий. Только первый переносчик протонов и электронов - никотинамидная дегидрогеназа расположена в матриксе митохондрии. Этот фермент отнимает водород от субстрата и передает его следующему переносчику. Полный комплекс таких ферментов образует «дыхательную цепь», в пределах которого атомы водорода отнимаются от субстрата, затем передаются последовательно от одного переносчика к другому, и, наконец, передаются на кислород воздуха с образованием воды. Существует строгая последовательность работы каждого звена в цепочке переносчиков. Эта последовательность определяется величиной редокс-потенциала (окислительно-восстановительного потенциала, сокращенно - овп) каждого звена. ОВП - это химическая характеристика способности вещества принимать и удерживать электроны. Вещества с положительным ОВП окисляют водород (отнимают от него электроны), вещества с отрицательным ОВП окисляются самим водородом. Самый низкий ОВП имеет начальное звено цепи, самый высокий - у кислорода, расположенного в конце цепочки переносчиков. Таким образом, передача водорода идет от более низкого к более высокому ОВП. На одной из стадий происходит разделение атомов водорода на Н+ и электроны. Протоны остаются временно в окружающей среде, а электроны идут дальше по цепи и в ее конце используются для активации О2. Кислород является конечным акцептором электронов. O2 + 4e -----> 2O-2 (полное восстановление кислорода). Все реакции, происходящие в дыхательной цепи, сопряжены. Переносчики водорода и электронов расположены в строгом порядке, в соответствии с величиной их редокс-потенциала. В настоящее время различают три варианта дыхательных цепей: Главная дыхательная цепь - это три мультиферментных комплекса, встроенных во внутреннюю мембрану митохондрии. Обозначаются они латинскими цифрами – I, III и IV. Сокращенная (укороченная) дыхательная цепи. Известны два субстрата. Главным из них является сукцинат (янтарная кислота). Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи - исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. DmH+ также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования - коэффициент Р/О=2. Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты): Максимально сокращенная (максимально укороченная) дыхательная цепь. Она представлена только цитохромной частью. Эксперименты показали, что здесь может быть окислен только один субстрат - аскорбиновая кислота, с участием фермента, восстановленные эквиваленты включаются в цепь на уровне цитохрома С цитохромоксидазы (цитохром аа3), но в реальных условиях такого окисления практически не происходит. Образуется вода и 1 молекула АТФ. Коэффициент Р/О=1.
44. Окислительное фосфорилирование, его сопряжение с дыхательной цепью. Химиоосмотическая теория Митчела. Коэффициент Р/О. Разобщение тканевого дыхания и окислительного фосфорилирования. Биологические разобщители как регуляторы окислительного фосфорилирования. Нарушения энергетического обмена: гипоксические состояния.
Окислительное фосфорилирование и дыхательный контроль. Функция дыхательной цепи – утилизация восстановленных дыхательных переносчиков, образующихся в реакциях метаболического окисления субстратов (главным образом в цикле трикарбоновых кислот). Каждая окислительная реакция в соответствии с величиной высвобождаемой энергии ≪обслуживается≫ соответствующим дыхательным переносчиком: НАДФ, НАД или ФАД. Соответственно своим окислительно-восстановительным потенциалам эти соединения в восстановленной форме подключаются к дыхательной цепи. В дыхательной цепи происходит дискриминация протонов и электронов: в то время как протоны переносятся через мембрану, создавая ΔрН, электроны движутся по цепи переносчиков от убихинола к цитохромоксидазе, генерируя разность электрических потенциалов, необходимую для образования АТФ протонной АТФ-синтазой. Таким образом, тканевое дыхание ≪заряжает≫ митохондриальную мембрану, а окислительное фосфорилирование ≪разряжает≫ ее. Разность электрических потенциалов на митохондриальной мембране, создаваемая дыхательной цепью, которая выступает в качестве молекулярного проводника электронов, является движущей силой для образования АТФ и других видов полезной биологической энергии. Механизмы этих превращений описывает хемиосмотическая концепция превращения энергии в живых клетках. Она была выдвинута П. Митчеллом в 1960 г. для объяснения молекулярного механизма. Согласно хемиосмотической концепции, движение электронов по дыхательной цепи является источником энергии для транслокации протонов через митохондриальную мембрану. Возникающая при этом разность электрохимических потенциалов (ΔμH+) приводит в действие АТФ-синтазу, катализирующую реакцию
АДФ + Рi = АТФ.
В дыхательной цепи есть только 3 участка, где перенос электронов сопряжен с накоплением энергии, достаточным для образования АТФ, на других этапах возникающая разность потенциалов для этого процесса недостаточна. Максимальная величина коэффициента фосфорилирования, таким образом, составляет 3, если реакция окисления идет с участием НАД, и 2, если окисление субстрата протекает через флавиновые дегидрогеназы. Теоретически еще одну молекулу АТФ можно получить в трансгидрогеназной реакции (если процесс начинается с восстановленного НАДФ): Обычно в тканях восстановленный НАДФ используется в пластическом обмене, обеспечивая разнообразные синтетические процессы, так что равновесие трансгидрогеназной реакции сильно сдвинуто влево. Эффективность окислительного фосфорилирования в митохондриях определяется как отношение величины образовавшегося АТФ к поглощенному кислороду: АТФ/О или Р/О (коэффициент фосфорилирования). Экспериментально определяемые значения Р/О, как правило, оказываются меньше 3. Это свидетельствует о том, что процесс дыхания не полностью сопряжен с фосфорилированием. Действительно, окислительное фосфорилирование в отличие от субстратного не является процессом, в котором окисление жестко сопряжено с образованием макроэргов. Степень сопряжения зависит главным образом от целостности митохондриальной мембраны, сберегающей разность потенциалов, создаваемую транспортом электронов. По этой причине соединения, обеспечивающие протонную проводимость (как 2,4-динитрофенол), являются разобщителями.
45. Свободно-радикальное окисление. Токсичность кислорода: образование активных форм кислорода, их действие на липиды и другие вещества клетки. Механизм защиты: антиоксидантные системы
Свободнорадикальное окисление – важный и многогранный биохимический процесс превращений кислорода, липидов, нуклеиновых кислот, белков и других соединений под действием свободных радикалов, а перекисное окисление липидов (ПОЛ) – одно из его последствий. Свободные радикалами (СР) представляют собой соединения, имеющие неспаренный электрон на наружной орбите и обладающие высокой реакционной способностью. К числу первичных СР относятся супероксидный анион-радикал, окись азота, а вторичными СР являются гидроксильный радикал, синклетный кислород, перекись водорода, пероксинитрит. Образование СР тесно связано, с одной стороны, с появлением свободных электронов при нарушениях процессов окисления в дыхательной цепи, превращении ксантина, синтезе лейкотриенов и простогландинов. Эти реакции зависят от активности ксантиноксидазы, дегидроротатдегидрогеназы, льдегидоксидазы, холестериноксидазы, ферментов цитохрома Р-450. Синтез супероксид — аниона инициируется ангиотензином II, который образуется из ангиотензина I под действием ангиотензинпревращающего фермента. Его активность в сосудах больных с выраженным атеросклерозом особенно высока. Причина этого явления пока неясна. Оксид азота образуется при окисленииL-аргинина под действием NO-синтетазы при участии кальмодулина. В литературе представлены данные о том, что синтез NO при атеросклерозе не нарушен, но его дефицит может возникать при увеличении его соединения с супероксид – анионом. Оно завершается синтезом пероксинитрита с большими патогенными потенциями, в том числе и образованием окисленных форм ЛПНП. С другой стороны, для синтеза вторичных СР используется водород НАДФ-Н, НАД-Н — его донаторов. Супероксид — анион может восстанавливать Fe3+ в Fe2+, при взаимодействии которого с перекисью водорода, перекисями липидов и гипохлоритом образуются высокотоксичные вторичные радикалы. Из всех СР наибольшей активностью обладают гидроксильный радикал и пероксинитрит. Активность СР ограничивается антиоксидантами, которые разрывают цепи молекул при реакциях СРО, разрушают молекулы перекисей. К числу ферментных антиоксидантов относятся супероксиддисмутаза (СОД), глютатионпероксидаза, каталаза, находящиеся в клеточных структурах. Неферментные антиоксиданты – витамины Е, К, С, убихиноны, триптофан, фенилаланин, церулоплазмин, трансферрин, гаптоглобин, глюкоза, каротиноиды – блокируют активность СР в крови.
46. Микросомальное окисление. Структурные компоненты и функции оксидаз, монооксигеназ и диоксигеназ. Значение микросомального окисления для организма: обезвреживание природных и искусственных экотоксикантов.
Свободное окисление. Одна из задач свободного (несопряженного) окисления – превращения природных или неприродных субстратов, называемых в этом случае ксенобиотиками (ксено – несовместимый, биос – жизнь). Они осуществляются ферментами диоксигеназами и монооксигеназами. Окисление протекает при участии специализированных цитохромов, локализованных чаще всего в эндоплазматическом ретикулуме, поэтому иногда этот процесс называют микросомальным окислением. В реакциях свободного окисления участвуют также кислород и восстановленные дыхательные переносчики (чаще всего НАДФН). Акцептором электронов является цитохром Р-450 (иногда цитохром b5). Окисление субстрата протекает по следующей схеме:
SH + O2 –> SOH.
Механизм действия оксигеназ включает изменение валентности входящих в их состав ионов двухвалентных металлов (железа или меди). Диоксигеназы присоединяют к субстрату молекулярный кислород, активируя его за счет электрона атома железа в активном центре (железо при этом становится трехвалентным). Оксигенация протекает как атака субстрата образующимся супероксид-анионом кислорода. Одной из биологически важных реакций такого типа является превращение β-каротина в витамин А. Монооксигеназы требуют участия в реакции НАДФН, атомы водорода которого взаимодействуют с одним из атомов кислорода, поскольку только один электрон связывается с субстратом. К широко распространенным монооксигеназам относятся разнообразные гидроксилазы. Они принимают участие в окислении аминокислот, оксикислот, полиизопреноидов. В процессе свободного окисления вследствие особенностей используемых цепей передачи электронов не происходит образования АТФ; биологическая роль этих процессов заключается в метаболизме ряда природных и ксенобиотических субстратов. В последнем случае свободное окисление выполняет важную функцию модификации чужеродных соединений.
47. Основные углеводы пищи. Переваривание углеводов в ротовой полости и желудочно-кишечном тракте, амилолитические ферменты слюны, поджелудочной железы, гидролиз дисахаридов. Всасывание моносахаридов (механизм всасывания).
По количеству углеводных остатков УВ разделяются на 3 основных класса: 1. моносахариды (глюкоза, фруктоза, галактоза, манноза и др.); 2. дисахариды (мальтоза, сахароза, лактоза); 3. полисахариды (гомополисахариды крахмал, гликоген, клетчатка). Крахмал – резервный гомополисахарид растений, построенный из остатков α-глюкозы. Крахмал представляет собой смесь двух гомополисахаридов: амилозы и амилопектина. В амилозе остатки глюкозы связаны α-1,4-гликозидными связями, в точках ветвления амилопектина - α-1,6-гликозидными связями. Молекулярная масса крахмала – 106-107. Гликоген – резервный гомополисахарид высших животных и человека, построенный из остатков α–Д-глюкозы. Гликоген содержится практически во всех органах и тканях человека и животных; наибольшее его количество находится в печени и мышцах. Молекулярная масса гликогена 107 – 109 и выше. Гликоген по своему строению близок к амилопектину. Остатки глюкозы связаны α-1,4-гликозидными и α-1,6-гликозидными связями (в точках ветвления). В молекуле гликогена имеется большее число α-1,6-гликозидных связей по сравнению с крахмалом. Клетчатка – это единственный гомополисахарид, который не переваривается в ЖКТ человека, т.к. пищеварительные железы человека не вырабатывают β–глюкозидазу. Однако клетчатка выполняет ряд важных функций: 1. способствует формированию кала; 2. усиливает перистальтику кишечника; 3. является адсорбентом, с которым из кишечника выводятся излишки холестерина, соли тяжелых металлов. Роль углеводов: 1. Энергетическая (глюкоза – основной источник энергии для организма. При сгорании 1 г УВ выделяется 4 ккал энергии). 2. Структурно-функциональная (УВ – обязательный компонент гликопротеинов и протеогликанов, которые выполняют разнообразные функции: гормональную, рецепторную, защитную, ферментативную и др.). 3. Метаболическая (пентозы участвуют в синтезе нуклеиновых кислот и нуклеотидных коферментов). Суточная потребность в углеводах составляет 400-500 г. Переваривание углеводов - это совокупность процессов поэтапного ферментативного гидролиза полисахаридов до моносахаридов, которые всасываются в кишечнике, разносятся током крови к печени и другим тканям организма, где подвергаются различным метаболическим превращениям. Переваривание УВ начинается в ротовой полости под действием фермента амилазы слюны (оптимум рН=6,8-7,2), которая гидролизует в крахмале α -1,4-гликозидные связи с образованием декстринов. Всасывание УВ из просвета кишечника в энтероцит и из энтероцита в кровь происходит: 1) облегченной диффузией с участием переносчиков; 2) вторично активным транспортом (симпортом с ионами натрия) с использованием энергии K, Na-АТФ-азы. Быстрее всех всасываются глюкоза и галактоза. От кишечника всосавшиеся моносахариды транспортируются в печень, где происходит до 90% превращений моносахаров. Поступление глюкозы в клетки сердечной, скелетных мышц и жировой ткани регулируется инсулином.
48. Потребность ребенка в углеводах. Своеобразие переваривания и всасывания углеводов в желудочно-кишечном тракте у ребенка.
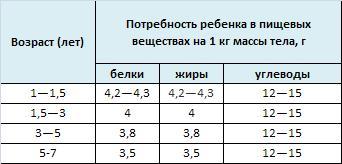
49. Обмен галактозы, фруктозы, маннозы. Взаимопревращение моносахаров в организме. Наследственные нарушения обмена моносахаридов: галактоземия, непереносимость фруктозы.
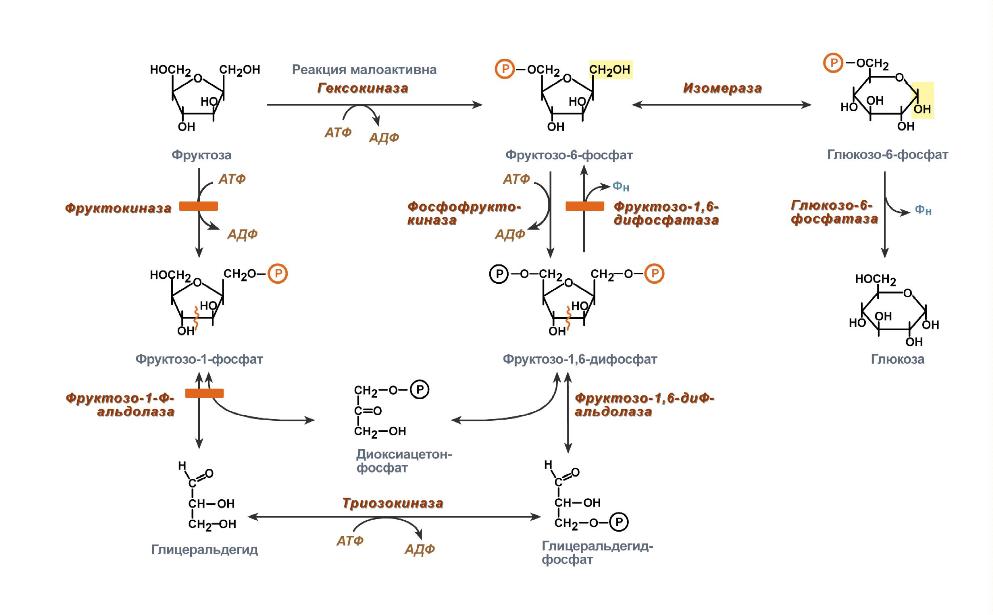
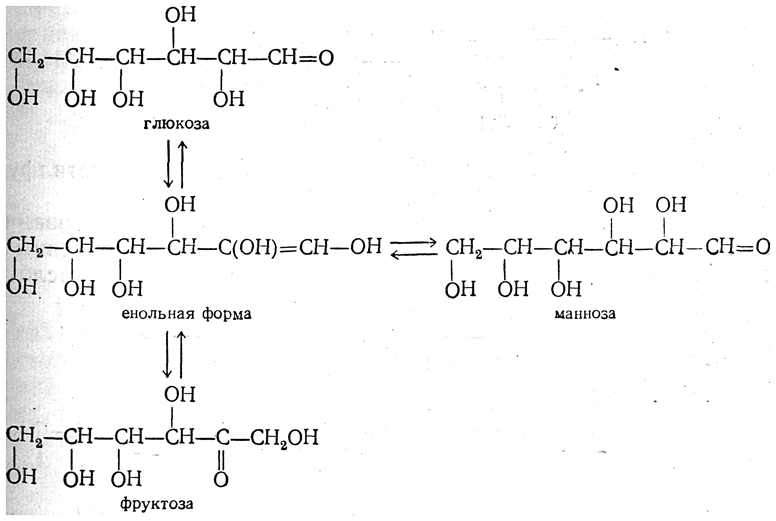
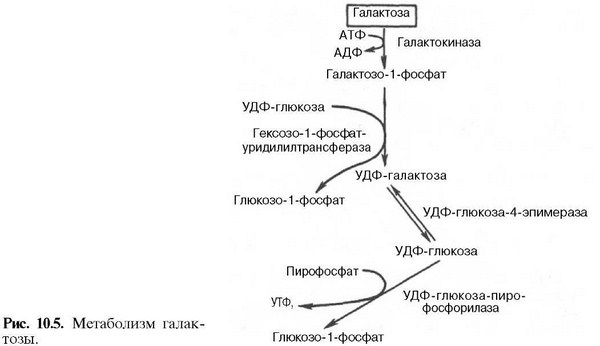
Нарушения, связанные с недостаточностью ферментов, которые принимают участие в переваривании и всасывании УВ. 1. Непереносимость лактозы: а) врожденная. Дефект по лактазе в просвете тонкого кишечника. Большой осмотический эффект невсосавшейся лактозы вызывает приток жидкости в тонкий кишечник, поэтому клиническими симптомами являются вздутие живота, тошнота, судороги, боль и водная диарея. Наследуется по аутосомно-рецессивному типу. б) приобретенная (временная). Может быть, и у взрослых вследствие инфекционных заболеваний или интенсивного лечения антибактериальными препаратами. 2. Нарушения обмена фруктозы: а) Фруктоземия (наследственная непереносимость фруктозы). Недостаточность фермента кетозо-1- фосфат-альдолазы приводит к резкому увеличению концентрации фруктозо-1-фосфата в клетках печени, вследствие чего развиваются хроническая недостаточность функций печени и почек, гипогликемия, диарея, рвота, боли в животе. б) Эссенциальная фруктозурия. Причина: недостаток фруктокиназы. Следствие: нарушение фосфорилирования фруктозы приводит к повышению содержания фруктозы в крови и аномальному выведению фруктозы с мочой. Это нарушение не вызывает патологических симптомов. 3. Нарушения обмена галактозы: а) Галактоземия. Причина: недостаток фермента гексозо-1-фосфатуридилтрансфераза печени. Больные дети плохо растут, прием молока вызывает рвоту и понос. Происходит увеличение печени и желтуха. При этом заболевании увеличивается концентрация галактозы и галактозо-1-фосфата. Галактоземия сопровождается галактозурией. У детей галактоземия приводит к умственной отсталости и катаракте хрусталика. Определяющим диагностическим критерием служит отсутствие в эритроцитах гексозо-1-фосфатуридилтрансферазы. б) Недостаток галактокиназы. Происходит накопление галактозы и ее превращение в галактитол. Следствие: раннее развитие катаракты.
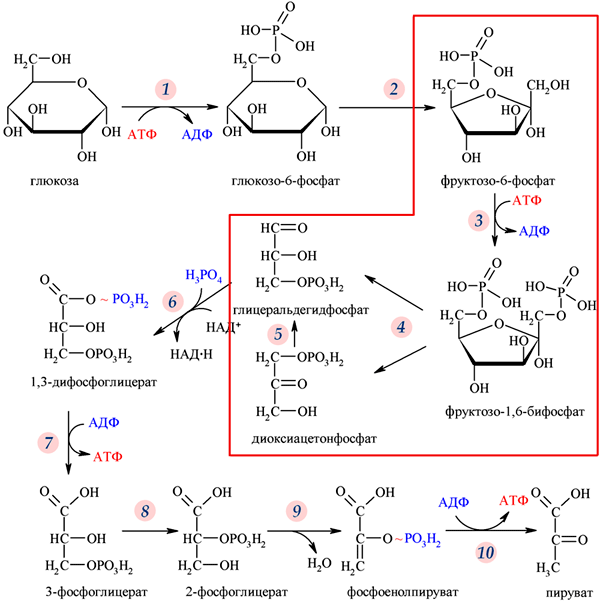 50. Анаэробный распад глюкозы. Этапы этого процесса. Гликолитическая оксиредукция, субстратное фосфорилирование. Энергетическая ценность анаэробного распада глюкозы. Регуляторные механизмы, участвующие в этом процессе.
50. Анаэробный распад глюкозы. Этапы этого процесса. Гликолитическая оксиредукция, субстратное фосфорилирование. Энергетическая ценность анаэробного распада глюкозы. Регуляторные механизмы, участвующие в этом процессе.
Гликолиз – синоним молочнокислого брожения – сложный ферментативный процесс превращения глюкозы до двух молекул молочной кислоты, протекающий в тканях человека и животных без потребления кислорода. Гликолиз включает 11 ферментативных реакций, протекающих в цитоплазме клетки. Реакции гликолиза проходят в 2 стадии. В ходе первой стадии – энергопотребляющей – используются 2 АТФ в 1-ой и 3-ей реакциях. В процессе 7-ой и 10-ой реакций второй стадии – энергодающей – образуются 4 АТФ. Из 11 реакций - 3 необратимые (1-ая, 3-я и 10-ая).
51. Анаэробный гликолиз, повышенная активность анаэробного распада глюкозы у ребенка раннего возраста. Биохимические критерии гликолиза. Связь с возникновением ацидоза.
Значение анаэробного гликолиза: 1. каждая молекула глюкозы в анаэробных условиях снабжает клетку 2-мя молекулами АТФ; 2. промежуточные метаболиты гликолиза (фосфоглицериновый альдегид, фосфодиоксиацетон, фосфоглицерат, пируват) могут использоваться клеткой в обмене липидов и белков; 3. пируват и НАДН используются в реакциях аэробного окисления глюкозы; 4. восемь реакций гликолиза из 11 обратимые, поэтому возможен синтез глюкозы из лактата (глюконеогенез). Эффект Пастера. Подавление анаэробного гликолиза аэробным окислением глюкозы. Факультативные анаэробы потребляют больше глюкозы, т.к. энергоотдача гликолиза – всего 2 АТФ. При добавлении О2 в среду потребление глюкозы резко уменьшается, т.к. возрастает количество АТФ, которое блокирует активность гексокиназы и фосфофруктокиназы. Спиртовое брожение. В процессе спиртового брожения молекула глюкозы в анаэробных условиях превращается в пируват, который затем декарбоксилируется и восстанавливается при участии НАДН с образование этанола. Особенности гликолиза и его нарушения у детей. У новорожденного аэробные процессы преобладают над аэробными. Это продолжается в течение первого месяца, пока фетальный Hb F не заменится на взрослый HbА. У детей высокая скорость аэробных процессов, в ходе которых вырабатывается АТФ, необходимая для синтетических процессов и роста. Детям требуется больше УВ в пище в качестве источника энергии. В детском организме анаэробные механизмы распада глюкозы и получения энергии подключаются при стрессах быстрее, чем у взрослых. Это свидетельствует о лучшей адаптации организма к экстремальным ситуациям. В подростковом возрасте еще не совершенны процессы гормональной регуляции (действие инсулина, глюкагона, адреналина). Распад гликогена может идти быстро, расход глюкозы большой. Но если не восполняется количество глюкозы в крови, легко наступает гипогликемия.
52. Аэробный распад глюкозы: этапы этого процесса и его энергетика. Физиологическое значение аэробного распада глюкозы.
Аэробный путь окисления глюкозы и его энергетика. Аэробный путь окисления глюкозы происходит в 3 этапа: 1. окисление молекулы глюкозы до 2 молекул пирувата; 2. окислительное декарбоксилирование пирувата (с образованием 2 молекул ацетил-КоА); 3. реакции цикла Кребса, где каждая молекула ацетил-КоА сгорает с образованием 12 АТФ. 1-ый этап. Окисление молекулы глюкозы до 2 молекул пирувата - это первые 10 реакций гликолиза. Суммарная реакция 1 этапа:
Глюкоза + 2 НАД++ 2 АДФ → 2 пирувата + 2 НАДН + Н+ + 2 АТФ
2-ой этап. Окислительное декарбоксилирование пирувата (с образованием 2 молекул ацетилКоА) происходит в матриксе митохондрий и осуществляется мультиферментным комплексом пируватдегидрогеназой. В его состав входят 3 фермента и 5 коферментов: 1. Пируватдекарбоксилаза – кофермент тиаминпирофосфат (активная форма витамина В1). 2. Дегидролипоилацетилтрансфераза – кофермента: в активном центре – липоевая кислота (витаминоподобное вещество), из матрикса митохондрий используется Коэнзим А (КоА~SH). 3. Дигидролипоилдегидрогеназа – коферменты: в активном центре – ФАД, из матрикса митохондрий используется – НАД. Суммарная реакция:
2 Пируват + 2 НАД+ + КоА → 2 ацетил-КоА + 2 НАДН + Н+
Каждый восстановленный НАД в митохондриях отдает протоны и электроны в полную ц.п.э., где образуется 2 х 3 АТФ = 6 АТФ. Таким образом, ценность второго этапа – 6 АТФ. Эти реакции происходят только в аэробных условиях! 3-ий этап. Реакции цикла Кребса, где каждая молекула ацетил-КоА сгорает с образованием 12 АТФ. Следовательно, энергетический выход третьего этапа (в расчете на молекулу глюкозы) – 2 х 12 АТФ = 24 АТФ . Энергетическая ценность аэробного окисления глюкозы: 1 этап – 8 (6) АТФ 2 этап – 6 АТФ 3 этап – 24 АТФ Итого: 38 (36) АТФ (разница в 2 АТФ зависит от челнока, который переносит протоны и электроны из цитозоля от НАДН, образованного в 6-ой реакции гликолиза в митохондрии в ц.п.э.). Аэробное окисление глюкозы является главным источником энергии для большинства клеток организма.
53. Интенсификация процесса прямого окисления глюкозы у детей раннего возраста. Причины этого явления.
54. Пентозофосфатный путь превращения глюкозы. Этапы и стадии этого процесса. Окислительная стадия пентозного цикла. Значение пентозофосфатного пути превращения глюкозы для организма.
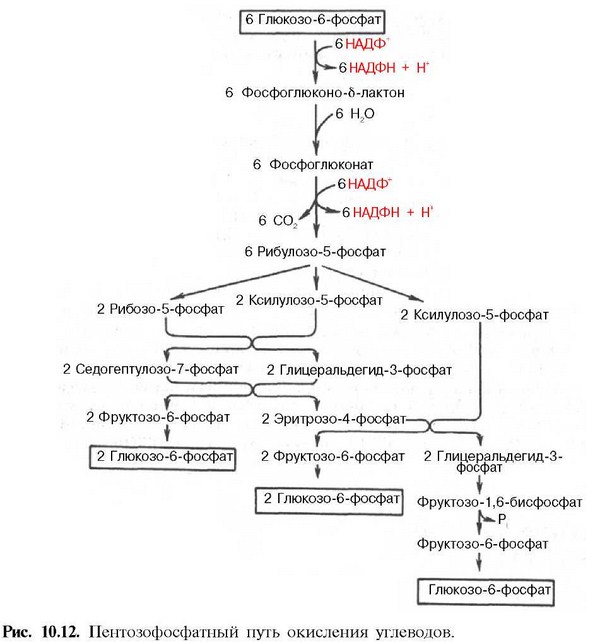
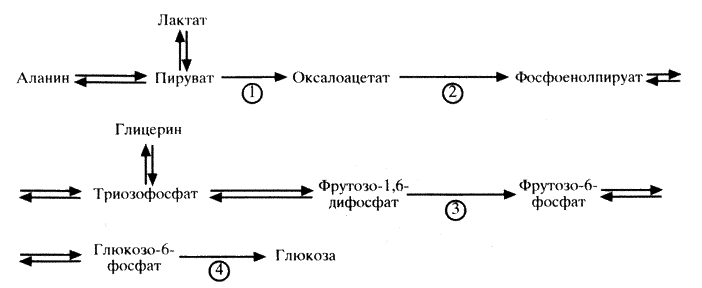
55. Биосинтез глюкозы (глюконеогенез) из неуглеводных продуктов. Основные этапы, три обходных реакции гликолиза. Значение процесса. Гормональная регуляция глюконеогенеза. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори).
Биосинтез глюкозы (глюконеогенез) из молочной кислоты. Глюконеогенез – обращение реакций гликолиза. Из 11 реакций гликолиза 3 необратимы (1-ая, 3-я и 10-я):
1. глюкоза → глюкозо-6 фосфат
2. фруктозо-6-фосфат → фруктозо-1,6-дифосфат
3. фосфоенолпируват → пируват
Для этих реакций существуют обходные пути. Первый обходной путь начинается в матриксе митохондрий. Пируват проникает сквозь мембрану митохондрий в матрикс митохондрии, затем пируват подвергается карбоксилированию под действием фермента пируваткарбоксилаза (с использованием биотина и энергии АТФ). В результате этой реакции образуется оксалоацетат. Затем оксалоацетат превращается в малат, чтобы выйти из митохондрий в цитозоль клетки для дальнейшего превращения. В цитозоле малат обратимо превращается в оксалоацетат (под действием цитозольной малатдегидрогеназы). Образовавшийся оксалоацетат под действием фермента фосфоенолпируваткарбоксикиназы (при участии биотина и энергии ГТФ) превращается в фосфоенолпируват. В дальнейшем реакции глюконеогенеза идут до образования фруктозо-1,6-дифосфата (обратимые реакции гликолиза). Второй обходной путь: Фруктозо-1,6-дифосфат + Н2О → фруктозо-6-фосфат + Фн (реакцию катализирует фруктозо-1,6-бифосфатаза) Третий обходной путь: Глюкозо-6-фосфат + Н2О → глюкоза + Фн (реакцию катализирует глюкозо-6-фосфатаза) В процессе глюконеогенеза 7-я реакция гликолиза (1,3-дифосфоглицерат + АДФ ↔ 3- фосфоглицерат + АТФ) расходуется 2 АТФ. В ходе синтеза глюкозы используются макроэргические соединения: 4 АТФ и 2 ГТФ. Регуляция глюконеогенеза происходит с участием гормонов и с участием метаболитов.
1. Гормональная регуляция. Глюкокортикоиды проникают в клетку, усиливают биосинтез ферментов глюконеогенеза, способствуют повышению уровня сахара в крови.
2. Метаболическая регуляция. Ацетил-КоА стимулирует пируваткарбоксилазу, осуществляющую реакцию превращения пирувата в ЩУК и далее в глюкозу. АТФ участвует в процессах карбоксилирования, стимулирует глюкозо-6-фосфатазу и фруктозо-1,6-бисфосфатазу, усиливает процессы глюконеогенеза. Витамин Н (биотин) – кофермент пируваткарбоксилазы. Фруктозо-2,6- дифосфат ингибирует фруктозо-1,6-бисфосфатазу.
Цикл Кори. В процессе анаэробного гликолиза в скелетных мышцах образуется лактат, который выходит из клеток в кровь, доставляется в печень, где включается в реакции глюконеогенеза с образованием глюкозы. Глюкоза выходит в кровь, доставляется к тканям, где используется как источник энергии. Цикл Кори выполняет две важные функции: 1) обеспечивает утилизацию лактата в реакциях глюконеогеназа; 2) предотвращает накопление лактата и развитие лактоацидоза (смещение рН в кислую сторону)
56. Глюконеогенез у детей. Лабильность этого процесса у ребенка.
57. Свойства и распространение гликогена как резервного полисахарида. Биосинтез гликогена, мобилизация гликогена. Регуляция обмена гликогена гормонами и аллостерическая регуляция. Особенности распада гликогена в печени и мышцах. Гликогенозы и агликогенозы
Гликоген – высокомолекулярный полимер, построенный из остатков глюкозы, связанных α-1,4- и α-1,6-гликозидными связями. В большом количестве содержится в печени и мышцах. Гликоген упакован в гранулы, размер которых 100 – 400 А. В гранулах находятся ферменты синтеза и распада гликогена. Распад гликогена происходит в интервалах между приемами пищи; этот процесс ускоряется при физических нагрузках. Гликогенолиз происходит за счет отщепления остатков глюкозы в виде глюкозо-1-фосфата. Фермент гликогенфосфорилаза расщепляет α-1,4-гликозидные связи в гликогене в результате остаются 4 остатка глюкозы до места ветвления (α-1,6-гликозидная связь). Олигосахаридтрансфераза переносит фрагмент гликогена (состоящий из трех остатков глюкозы) на неразветвленный участок цепи, остается один остаток глюкозы, связанный α-1,6-гликозидной связью. Фермент α-1,6-глюкозидаза отщепляют мономерный остаток глюкозы. Суммарная реакция: Глюкозо-1-фосфат изомеризуется в глюкозо-6-фосфат под действием фосфоглюкомутазы. Образующийся глюкозо-6-фосфат вступает в реакции гликолиза (или пентозного цикла) клеток мышц и мозга, а в клетках печени, почек и кишечника происходит дефосфорилирование глюкозо-6-фосфата с образованием глюкозы и фосфорной кислоты (под действием фермента глюкозо-6-фосфатазы). Далее глюкоза поступает в кровь и разносится с кровью к тканям. Гормональная регуляция обмена гликогена осуществляется адреналином, глюкагоном и инсулином. Адреналин и глюкагон активируют распад гликогена и тормозят его синтез. Гормональная регуляция опосредуется общими механизмами через ц-АМФ, который активирует цитозольные ферменты протеинкиназу, киназу фосфорилазы и киназу гликогенсинтазы. Киназы при участии АТФ фосфорилируют ферменты гликогенфосфорилазу и гликогенсинтазу. Фосфорилаза активна в фосфорилированном состоянии, поэтому под действием адреналина и глюкагона активируется распад гликогена, а гликогенсинтаза, напротив, в фосфорилированном состоянии становится неактивной, поэтому синтез гликогена под действием адреналина и глюкагона тормозится. Под действием инсулина активируются ферменты фосфодиэстераза и фосфопротеинфосфатазы. Фосфодиэстераза разрушает ц-АМФ; фосфопротеинфосфатазы катализируют дефосфорилирование ферментов гликогенфосфорилазы и гликогенсинтазы. Фосфорилаза в дефосфорилированном состоянии становится неактивной и процесс распада гликогена прекращается, а гликогенсинтаза, напротив, активируется, поэтому под действием инсулина активируется синтез гликогена. Заболевания, связанные с обменом гликогена: гликогенозы и агликогенозы. 1. Болезнь Гирке (недостаток глюкозо-6-фосфатазы печени). Клиническая картина: увеличение печени уже в период новорожденности, гипогликемия между приемами пищи, гипогликемия, сохраняющаяся при действии адреналина и глюкагона, нарушение жирового обмена (усиливается отложение жира в подкожной клетчатке и внутренних органах). 2. Болезнь Форбса-Кори (дефицит фермента амило-1,6-глюкозидазы печени): неполное расщепление гликогена, гепатомегалия, мышечная слабость, гипогликемия. Эта болезнь не угрожает жизни ребенка, хотя может привести к задержке физического развития и полового созревания. У некоторых больных в подростковом возрасте отмечается умеренная миопатия, т.к. понижена мышечная утилизация гликогена. 3. Болезнь Андерсена (недостаток активности «ветвящего» фермента в печени): гепатоспленомегалия, цирроз печени, прогрессирующая печеночная недостаточность - длинные неразветвленные цепи плохо разрушаются; развивается цирроз печени. Эта болезнь встречается крайне редко и всегда приводит к ранней смерти. 4. Болезнь Херса (недостаток активности печеночной фосфорилазы): гепатомегалия, умеренная гипергликемия, ацидоз. Данная патология встречается крайне редко и только у мальчиков. 5. Болезнь Мак-Ардля (недостаток мышечной фосфорилазы): спазмы мышц при нагрузке, преходящая миоглобинурия. Агликогенозы – наследственное заболевание, вызванное дефектом гликогенсинтазы. В печени почти или полностью отсутствует гликоген, выражена гипогликемия (0,39 – 0,67 ммоль/л). Характерный симптом – у детей судороги обычно по утрам. Их можно предупредить частым кормлением ночью. Гипогликемия новорожденных. Наблюдается у всех новорожденных из-за прекращения поступления крови через плаценту (преходящая гипогликемия). Коррекция происходит быстро и самостоятельно. У новорожденных, матери которых страдают сахарным диабетом, может быть тяжелая гипогликемия – проявление функционального гиперинсулинизма, устраняемое только введением глюкозы
58. Своеобразие метаболизма гликогена в печени у ребенка. Патология обмена гликогена (гликогенозы).
Биосинтез гликогена. При голодании гликоген распадается до небольшого фрагмента, содержащего 4-6 остатков глюкозы, связанных α-1,4-гликозидными связями. Этот фрагмент гликогена называется «затравкой» (или праймером); на нем начинается процесс биосинтеза гликогена. Глюкоза вступает в синтез гликогена в «активной» форме УДФ-глюкозы. В период пищеварения глюкоза поступает в клетку и фосфорилируется с образованием глюкозо-6-фосфата под действием фермента гексокиназы (или глюкокиназы в печени), затем изомеризуется в глюкозо-1-фосфат под действием фосфоглюкомутазы. Глюкозо-1-фосфат превращается в УДФ-глюкозу под действием фермента УДФ-глюкуронилтрансферазы. Реакция, катализируемая гликогенсинтазой, возможна лишь в том случае, когда в цепи «затравки» гликогена содержится более 4-6 остатков глюкозы: