
Антигены вирусов и преодоление иммунитета








Антигены вирусов – это белки и гликопротеиды их суперкапсида, капсида, внутренние белки-ферменты и нуклеопротеиды. Так, у вируса гриппа основными антигенами служат нейтроаминидаза и гемагглютинин, у вируса гепатита В – поверхностный НВS антиген, а также НВе, НВС, у ВИЧ вируса – его белки р14, 18 и гликопротеиды – gр120 и другие. У вируса гепатита А идентифицировано более 40 антигенореактивных доменов в структурных и неструктурных белках. Каждая такая антигенная молекула имеет много антигенных эпитопов, поэтому антитела к ним могут отличаться по специфичности. Кроме того, антигенная структура многих вирусов может изменяться, что препятствует развитию иммунитета. Протективными свойствами – способностью индуцировать иммунитет обладают поверхностные, оболочечные антигены вирусов.
Вирусы уклоняются от элиминации системой иммунитета, изменяя антигенные свойства. Точечные мутации вызывают небольшие изменения (антигенный дрейф), а большие изменения, приводящие к эпидемиям, могут возникать в результате пересортировки сегментов генома или обмена генетическим материалом с другими вирусами, имеющими иных хозяев (антигенный шифт).
Зараженные вирусом клетки экспрессируют на своей поверхности его антигены, так как оболочки вирусов часто формируются из клеточных мембран. Если экспрессируется белок слияния, то клетки образуют синцитий. Вирусные антигены на поверхности клеток распознаются системой иммунитета с образованием антител и Т-киллеров. Антитела и Т-киллеры специфичны против разных эпитопов одного антигена.
Иммунитет возникает если уничтожаются свободные вирионы или/и зараженные ими клетки.
Вирусные антигены (наряду с антителами) могут присутствовать в крови и других биологических жидкостях больных. Их выявление (обычно методом ИФА или РИФ) используется для диагностики инфекций.
Приобретенный противовирусный иммунитет
Резистентность к вирусам в иммунном организме, например, после вакцинации вирусными вакцинами, при прочих равных условиях с неиммунным организмом по неспецифической резистентности, зависит от наличия специфических факторов иммунитета – IgG, IgM, секреторных IgA антител, возможно IgD антител, а также иммунных Т-киллеров.
Все вирусные антигены являются Т-зависимыми. Антигенпредставляющие клетки презентируют одни вирусные антигены, связанные с HLA I класса, CD8+ Т-лимфоцитам, из которых возникают иммунные Т-киллеры. Другие антигены представляются в комплексе с HLA II класса CD4+ Т-хелперам, которые индуцируют синтез антител к вирусным антигенам вначале IgM, а затем IgG-класса. Антитела против вирусных антигенов, даже в низких концентрациях, способны нейтрализовать вирус, блокируя его рецепторы и проникновение через входные ворота в кровь и/или фиксацию на клетках-мишенях (IgG, IgM), а также при первичном попадании его на эпителий слизистых – sIgA может связывать их даже в эпителиальных клетках. Это объясняет высокую эффективность вакцинации при долговременной профилактике и эффективность введения специфических иммуноглобулинов для экстренной кратковременной профилактики при многих вирусных инфекциях. Антитела, при их наличии в достаточном количестве, могут нейтрализовать свободные вирионы, особенно в тех случаях, если они находятся в крови внеклеточно. Однако антитела только блокируют вирионы, а их лизис осуществляют компоненты активированного комплемента. Разрушать вирион, «покрытый» антителами, могут клетки, осуществляющие антителозависимую клеточную цитотоксичность. Антитела же обеспечивают защиту и от повторного заражения. Они эффективны при кори, полиомиелите, паротите, краснухе, гриппе (к конкретному серотипу) и других инфекциях. При таких инфекциях уровень антител отражает напряженность иммунитета. Однако антитела не всегда эффективны против вирусов, особенно после их проникновения в клетку.
Появление антител у больных не ликвидирует развившуюся ВИЧ-инфекцию, гепатиты и другие инфекции. Для этого необходимо дополнительное сочетание факторов: активированные макрофаги, Т-киллеры, активация интерферонами резистентности к вирусам у клеток-мишеней. В некоторых ситуациях антительный иммунный ответ препятствует развитию эффективного Т-клеточного ответа (конкуренция активности Тх 2 и Тх 1). Более того, покрывая вирус, но не повреждая его, антитела могут усиливать его проникновение в клетку, связываясь своими Fc-фрагментами с Fc-рецепторами клеток (например, вирус денге).
Вирусы, которые проникают в соседние клетки, минуя встречу с антителами, уничтожаются механизмами клеточного иммунитета. Макрофаги фагоцитируют вирусы, и многие из них разрушают. Фагоцитоз усиливается, если вирион опсонирован антителами. Однако некоторые вирусы, например ВИЧ, резко активируют макрофаги, которые выделяют избыток цитокинов (ИЛ-1, ФНОa), повреждающих другие клетки, но не вирусы.
Важным фактором противовирусного иммунитета служат вирусспецифические Т-киллеры.
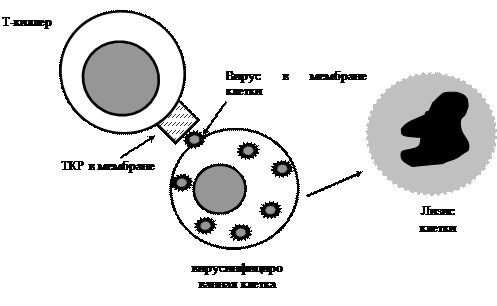
При большинстве контролируемых вирусных инфекций Т-клетки либо элиминируют вирус, либо супрессируют его, что приводит к развитию безвредной персистентной инфекции. Однако, например, ВИЧ инфицирует ключевые клетки СИ – CD4+ и дезорганизует ее реакции. Инфицированные клетки начинают экспрессировать поверхностные вирусные антигены через короткое время после проникновения в них вируса. Быстрое уничтожение таких клеток цитотоксическими Т-лимфоцитами (рис. 3.2) предотвращает репликацию вируса, а Т-хелперы 1 типа, выделяя гамма-интерферон, подавляют репликацию вируса в здоровых клетках. Вирус-специфические Т-клетки находят как при иммунитете, так и при персистирующей инфекции, однако для иммунитета количество их должно быть достаточным.
 |
ТКР – рецептор Т-киллера