
Варианты приобретенного антибактериального иммунитета








Приобретенный иммунитет к бактериальным инфекциям различается по механизмам в зависимости от факторов патогенности возбудителя. В одних случаях, когда бактерии выделяют токсины, или чувствительны к антителам, он эффективен, в других – неэффективен, например, при индукции антител к внутриклеточным бактериям, в третьих – при выделении избытка цитокинов, иммунный ответ повреждает собственные ткани.
Бактериальные инфекции, которые зависят от продукции экзотоксинов, индуцируют антитоксический иммунитет (дифтерия, столбняк, ботулизм и др.). Ведущая роль в нейтрализации токсинов принадлежит IgM- и IgG-антителам. IgM-антитела в крови выявляются уже через 48 часов после заражения и достигают пика через 7-10 дней (при инфекциях – позже). Затем преобладают IgG-антитела. Молекула антитела, присоединившись вблизи активного центра токсина, может стереохимически блокировать его связь с рецептором. В комплексе с антителами токсин теряет способность к диффузии в тканях и может стать объектом фагоцитоза.
Основным механизмом антибактериальной защиты является фагоцитоз (рис. 3.1). В иммунном организме эффективность фагоцитоза повышается за счет опсонизирующего действия специфических IgM- и IgG-антител, взаимодействующих Fab-фрагментами с антигенами на поверхности бактерий и одновременно с Fc-рецепторами на мембранах фагоцитов. Это приводит к окислительному взрыву и активации других бактерицидных систем фагоцитирующих клеток.
Активация системы комплемента комплексами «антитела-бактерии» приводит к разрушению липопротеиновых оболочек грамотрицательных бактерий, особенно нейссерий, а также к высвобождению анафилотоксинов, которые стимулируют дополнительный приток из плазмы крови гуморальных компонентов

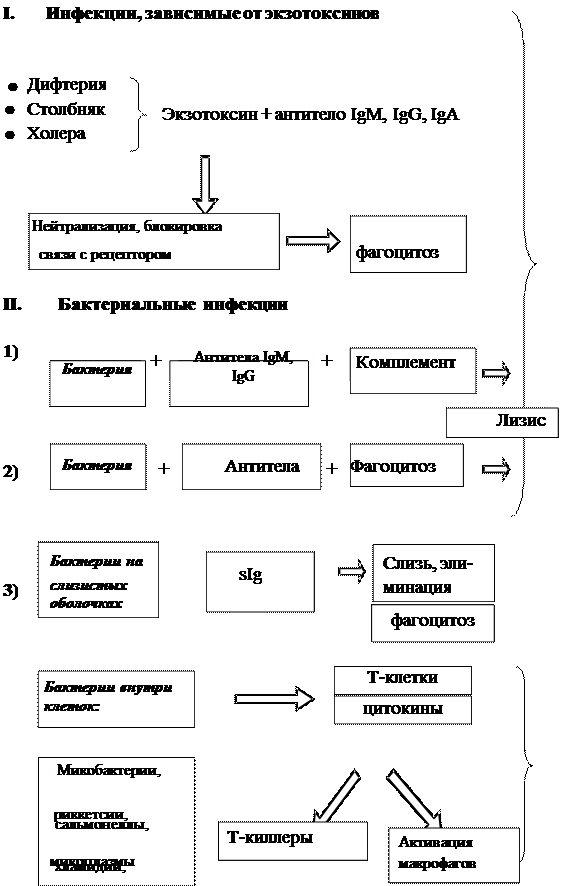
Рис. 3.1. Особенности антибактериального иммунитета
иммунитета и вызывают хемотаксис полиморфноядерных лейкоцитов, осуществляющих фагоцитоз.
Некоторые бактерии уклоняются от контактов с фагоцитирующими клетками, прикрепляясь к поверхности слизистых оболочек и заселяя их. Функцию защиты слизистых оболочек выполняет секреторный IgA. Во всех секретах IgA, связавшись с бактериями, предотвращает их адгезию к поверхности слизистой.
Приобретенный антибактериальный иммунитет, особенно с антителами против полисахаридных антигенов, как правило, является типоспецифическим и нестойким. Этим объясняются частые случаи повторных заболеваний бактериальными инфекциями и необходимость проведения частых ревакцинаций при использовании бактериальных профилактических вакцин, формирование нестерильного иммунитета, или неэффективность вакцинации при отдельных бактериальных инфекциях.
Липополисахариды бактерий индуцируют синтез антител к полисахаридным детерминантам, которые не всегда являются «иммунодоминантными», так как такие антитела не создают иммунитет, что наблюдается при бруцеллезе, туберкулезе и других инфекциях. В то же время такие антитела эффективны против E . coli, лептоспир и других бактерий.
Внутриклеточно паразитирующие бактерии: микобактерии туберкулеза, бруцеллы, сальмонеллы и др., а также риккетсии, хламидии и микоплазмы отличаются повышенной устойчивостью к гибели после фагоцитоза. Они защищаются от механизмов уничтожения, подавляя слияние фагосом с лизосомами, образуя наружную оболочку, или выходя из фагосом в цитоплазму. Эти бактерии уничтожаются механизмами Т-клеточного иммунитета. Специфические цитокин-продуцирующие Т-хелперы при контакте с зараженными макрофагами выделяют g-интерферон, активирующий ЕК и макрофаги, которые становятся после этого более эффективными, что напоминает противовирусный иммунитет. Однако важнейший механизм – это индукция активности Т-киллеров, которые разрушают инфицированные клетки и делают доступными бактерии для других бактерицидных факторов, в том числе для активированных макрофагов.
Поэтому напряженность антибактериального иммунитета при внутриклеточных инфекциях определяется не гуморальным, а Т-клеточным иммунитетом. Причем разные Т-субпопуляции оказывают различный эффект: преобладание активности Тх 2 у людей способствует заболеванию туберкулезом. Среди цитотоксических эффекторов одни разрушают зараженные клетки хозяина путем апоптоза и выделения цитокинов, а другие лизируют непосредственно сами бактерии.
Выраженность и сила этого иммунитета определяется путем постановки кожно-аллергических проб и в тестах оценки Т-клеточного иммунитета in vitro.
В целом же защита от внутриклеточных бактерий, особенно после иммунизации, носит комбинированный характер и в ней участвуют активированные фагоциты, поглощающие опсонированные микробы. В большинстве случаев для оценки уровня противобактериального иммунитета применяют различные методы выявления антител в сыворотке крови, даже если их уровень и не определяет напряженности антибактериального иммунитета.
Для серологической диагностики используют феномен нарастания титра циркулирующих антител в динамике инфекционных заболеваний (метод парных сывороток), или определение в острую фазу заболевания ранних IgM-антител. IgG-антитела появляются несколько позже, в период ранней реконвалесценции, и могут циркулировать в течение всей жизни, как после перенесенного заболевания, так и после вакцинации.
Таким образом, иммунитет к бактериям формируется в результате постоянного взаимодействия между СИ организма и микробами, изменяющими свои свойства, эволюционная цель которых – выжить и противостоять действию этой системы. Выживаемость микробов обеспечивается защитой от фагоцитоза за счет капсул, секрецией экзотоксинов, подавляющих фагоциты и иммунные реакции. Иногда микробы заселяют относительно недоступные для СИ места организма. Антитела обеспечивают иммунитет, нейтрализуя токсины, активируя комплемент непосредственно на поверхности бактерий, преодолевая антифагоцитарные свойства капсулы, опсонируя её с помощью IgG и C3b. Недостаточный эффект антител могут дополнить Т-киллеры.