
2. Анохин П.К. Очерки по физиологии функциональных систем. – Москва, 1975 – 476 с.








Цуканов Б.И.
ЭВОЛЮЦИОННОЕ ЭХО В НАТУРАЛЬНОМ ПОРЯДКЕ ТИПОЛОГИЧЕСКИХ ГРУПП.
Первая дифференциация индивидуальных различий связывается с именем Гиппократа, который будто бы разделил людей на четыре типа темперамента по преобладанию одного из четырех соков в организме индивида [14]. Гиппократ сумел увидеть в разнообразии индивидов такие черты, которые позволили ему установить конечную группировку, “одинаково справедливую для представителей различных эпох, наций и сообществ” [16, с.12]. Сама реальность существования четырех типологических групп в природе человеческой популяции приводила к тому, что каждый из исследователей темперамента, закладывая в его основания те или иные типологические признаки (черты, свойства), неизбежно приходил к “собственному видению” конечного числа типов, которое в большинстве теоретических схем соответствовало четырем классическим типам, связанным с именем Гиппократа. На данный момент понятно, что Гиппократ говорил о наиболее ярких представителях типологических групп, которых, по степени выраженности черт и их соотношении можно считать “чистыми типами” Такой порядок имеет место в самой природе человеческой популяции, и именно в таком порядке впервые увидел размещение четырех классических типов “греческий гений в лице Гиппократа”, сумевший уловить “в массе бесчисленных вариантов человеческого поведения капитальные черты” [13, с.280].
Бытует мнение, что Гиппократ разделил людей на четыре типа по преобладанию одной из четырх “влаг” в организме, и именно по этому преобладанию дал названия темпераментам. Но, как свидетельствует история медицины [5], Гиппократ разделил людей на типы по “предпочитаемым” болезням. Наиважнейшим достижением Гиппократа является создание им учения о психосоматическом единстве индивида. Положив в основу дифференциации индивидов “предпочитаемые” болезни (объективный показатель), Гиппократ установил, что холерики по своим “предпочитаемым” болезням составляют полную противоположность флегматикам, а последние болеют только своими “предпочитаемыми” заболеваниями. Сангвиники болеют по типу холериков, а меланхолики – по типу флегматиков. Так был открыт последовательный линейный порядок расположения типов.
Необходимо отметить, что в проблеме дифференциации индивидуальных различий современные исследователи, имея дело с психосоматическим единством, в своих подходах пытались найти факторы, или системы, с помощью которых им удавалось установить и объяснить те различия, которые реально (подчеркнуто нами, - Б.Ц.) существуют у индивидов человеческой популяции. Как это ни досадно, но ни один из авторов теорий темперамента никогда не ставил вопрос: как могла быть создана именно такая природа индивидуальных различий в типологических группах и почему существует именно такой их порядок? Подобный вопрос впервые привлек внимание основоположника дифференциальной психологии Фр.Гальтона. [15]. К сожалению, другие исследователи стыдливо обходили этот принципиальный вопрос стороной и он оставался открытым.
Натуральный порядок типологических групп был восстановлен нами по линейному спектру “τ-типов”, который фактически представляет собой последовательную цепь индивидуальных квантов времени [19]. Поэтому вопрос о природе индивидуальных различий – это прежде всего вопрос о происхождении последовательной цепи квантов времени линейного спектра “τ-типов”. Ответ на последний вопрос был найден нами в механизме отсчета времени жизни Глобальными биологическими часами (ГБЧ) [23]. По закону дискретного хода в горниле ГБЧ был создан эукариот наивысшего уровня организации – человек современный. Поэтому ответ на вопрос о присхождении типологических групп в человеческой популяции, о их количестве и натуральном порядке следует искать в механизме дискретного отсчета ГБЧ собственного времени.
Универсальность индивидуальных различий общепризнанна потому, что они, как подчеркивала А. Анастази [1, с. 49], имеют место у всех видов животных, начиная с одноклеточных организмов и, заканчивая антропоидами. Такую же универсальность имеют и спонтанные околосуточные ритмы у эукариотов (17 час≤Сd≤30 час) всех уровней организации, начиная с одноклеточных и, заканчивая человеком современным [4]. Учитывая значение Cd-периодов в жизнедеятельности эукариотов и согласованность их продолжительности с замедлением вращения Земли около собственной оси, на 0,0015с. за 100 лет [23], начиная с момента T6=3,3K (появление эукариотов) на шкале времени жизни, вопрос о происхождении индивидуальных различий может быть сформулирован так: когда и в каких особенных точках на шкале времени жизни происходило последовательное отделение и разграничивание эукариотной биоты на типологические группы в соответствии с законом дискретного хода ГБЧ?
Известно, что мозг индивида выполняет роль центрального механизма в собственных биологических часах и несет ответственность за точность подстройки фазы Cd-периода к длительности суточного вращения Земли [18]. Механизм подстройки фазы, или механизм затягивания ритма [3], [4], [18] срабатывает в момент перехода от света к темноте (С→Т), или в момент перехода от темноты к свету (Т→С), чему в природных условиях соответствует смена дня ночью, или смена ночи днем. Синхронизация Cd-периода индивида с чередованием света и темноты в пределах суточного вращения Земли требует исключительной точности [18].
Околосуточный период жизнедеятельности индивида складывается из двух фаз: фазы активности (период бодрствования) и фазы покоя (период сна). В наших исследованиях был обнаружен высокий уровень корреляционной зависимости длительности фазы активности (r=+0,96) и длительности фазы покоя (r=-0,98) в пределах Cd–периода от длительности собственной единицы времени [22]. В линейном спектре “τ-типов” по длительности периода сна (Тс) выявлены следующие группы индивидов:
1) индивиды с Тс >10 час (0,7с ≤ τ < 0,8с);
2) индивиды с Тс >8 час (0,8с ≤ τ < 0,9с);
3) индивиды с Тс ≈ 8 час (τ ≈ 0,9с);
4) индивиды с Тс < 8 час (0,9с < τ ≤ 1,0с);
5) индивиды с Тс < 6 час (1,0с < τ ≤ 1,1с).
Как видно из приведенного, линейный спектр “τ-типов” розграничивается на четыре типологические группы в тех же критических точках( τ1=0,8с; τ2=0,9; τ3=1,0с), которые были обнаружены ранее методом наименьших квадратов [19]. Высокий коэффициент корреляции говорит о том, что собственная единица времени в биологических часах индивида представляет собой системообразующий квант длительности, в соответствии с которым Cd-период делится на две фазы в строгой пропорции. Обязательным условием подобной пропорции является то, что сумма фаз бодрствования и сна у индивидов всех типологических групп согласована с периодом теперешнего суточного вращения Земли с исключительной точностью. Установленная зависимость еще раз подтверждает универсальность кванта времени индивида, который выполняет функцию “шага” для измерения времени в его собственном Cd-периоде жизнедеятельности. На основе этой зависимости мы выдвинули предположение, что в линейном спектре “ τ – типов ” зафиксирована вся последовательность реальных квантов длительности, с помощью которых эукариоты, появившись в горниле ГБЧ, согласовывали свои Cd-периоды жизнедеятельности с замедлением суточного вращения Земли. Для проверки предположения можно использовать простое уравнение:
Cd = 24·τ (1).
Подставляя в это выражение квант τ = 0,7 легко получить Cd=16,8 час. Учитывая разницу ∆Cd можно убедиться, что суточное вращение Земли за 16,8 часа происходило на интервале Т7=1,728·109 лет от “нулевой” точки, за которую на современной геохронологической шкале Шопфа условно принят 1950 год [11]. На шкале времени ГБЧ это точка 3К, в окрестности которой по палеонтологическим данным следы эукариотов уже четко зафиксированы в истории Земли. Таким образом, в особенной точке 3К произошло выделение группы эукариотов, у которых квант согласования собственного Cd-периода с суточным вращением Земли равнялся 0,7с. Само собой разумеется, что эукариоты, выделившись в самостоятельную группу в структуре ГБЧ, для согласования собственных Cd-периодов с равномерным замедлением суточного вращения Земли, последовательно использовали кванты времени все большей длительности. Так, в горниле ГБЧ был выстроен линейный спектр последовательного увеличения квантов времени для согласования собственных Cd-периодов эукариотов. Эта последовательность и была обнаружена нами на репрезентативной выборке эукариотов – “человек современный” в виде линейного спектра “τ-типов” [22].
Теперь попробуем ответить на вопрос: когда произошло разграничение линейного спектра “τ-типов” на типологические группы? Подставляя в формулу (1) τ1=0,8, получим Cd=19,2 часа. За такой период суточное вращение Земли происходило на интервале Т8=1,15·109 лет. Это особенная точка 2К, в которой эукариоты резко переходят от ферментации к дыханию и половому размножению (известная “точка Пастера”).
Для кванта τ2=0,9 получим Cd = 21,6 часа. Именно за такой отрезок времени Земля совершала суточный оборот на интервале Т9=576·106 лет удаленности от “нулевой” точки. На шкале времени жизни это особенная точка К, в которой зафиксировано “взрывообразное” начало скелетной революции [11].
Очередной, с “точки зрения” ГБЧ, и наиболее особенный, с точки зрения нас, сознательных индивидов момент на шкале времени жизни, - это момент появления человека современного. От точки К до этого момента период суточного вращения Земли замедлился до 24-х часового, а квант согласования Cd-периода практически равен 1с. Говоря строго, с учетом исключительной точности синхронизации Cd-периодов с длительностью суточного вращения Земли, в момент появлеия кроманьйонца суточный период был всего на ΔCd=0,53с меньше нынешнего, а длительность кванта согласования составляла τ3=0,9999938с. Возраст человечества по подстройке фазы всего 0,53с! Не удивительно, что человечество выбрало именно последний квант длительности, созданный ГБЧ для подстройки к 24-х часовому суточному периоду, назвало его секундой и приняло как эталон для измерения времени. Исходя из изложенного можно утверждать, что размежевание линейного спектра “τ-типов” на четыре типологические группы – это своеобразное эволюционное эхо от трех особенных точек на геохронологической шкале времени жизни, в которых по закону дискретного хода ГБЧ были зафиксированы резкие скачки в биосфере [23].
Появившись в точке Т=3,3К, эукариоты, согласно [24] сначала мирно сосуществуют с прокариотами до точки собственного выделения (Т=3К). Начиная с этого момента эукариоты стают агрессивными по отношению к прокариотам и начинают активно их вытеснять, завоевывая пространство собственного существования [24]. Эта «война» продолжалась на протяжении интервала от 3К до 2К. Так были установлены пределы первой (холероидной) группы, у индивидов которой гневливость и открытая враждебность стали доминирующей эмоциональной окраской их поведения. В точке 2К эукариоты резко переходят от ферментации к дыханию и половому размножению. Именно эти новые элементы в их жизнедеятельности, вероятно, и стали основой для появления “эмоции удовольствия”. Вспомним, что доминирующие и ярко выраженные эмоции радости и удовольствия украшают поведение индивидов сангвиноидной группы и поныне.
Согласно палеонтологическим данным поблизости точки 2К жизнь на Земле была достаточно разнообразной [11]. За период от 2К до К происходит обособление групп беспозвоночных, появляются настоящие многоклеточные, а трехслойные животные разделяются на первичноротых и вторичноротых [9]. Все типы и классы вендской фауны (докембрийский период) появляются в палеонтологической летописи как бы внезапно. Но в Венде, как подчеркивает Мороз [11, с.84], отсутствуют активные хищники и трупоеды! Так и хочется думать, что в фауне того периода была «радостная и счастливая» жизнь, господствовали «мир и согласие».
Точка К знаменательна началом “взрывообразной” скелетной революции. С этого момента ГБЧ переходят на “ускоренный” ход. Особенные точки, в которых фиксировались те или иные “знаменательные” скачки в биосфере размещаются на шкале временни жизни с нарастающей плотностью. Как и когда, согласно закону дискретного хода ГБЧ, в посткембрии появились и исчезли те или иные представители флоры и фауны, детально изложено в нашем отдельном исследовании [23]. Отметим только, что главным «дирижером» оказались ГБЧ, которые разрешали тем или другим группам эукариотов существовать только в строгих пределах таких интервалов, которые соответствовали их способности к затягиванию Cd-периода. Выбравшие свой резерв группы эукариотов исчезали “внезапно”. Такая участь постигла многих представителей посткембрия: трилобитов, брахиопод, ракоскорпионов, кистеперых рыб, динозавров, аллотерий, древних хищников, слонов, коней, китов и многих других предшественников человека современного [23]. Фактически в посткембрии многократно разыгрывалась драма (если не трагедия), которая порождала у эукариотов эмоцию страха перед будущим, ибо «перспектива» внезапно исчезнуть определялась только по закону дискретного хода ГБЧ. Довлеющая эмоция страха перед будущим у индивидов меланхолоидной группы и поныне остается тем свойством, которое резко отличает их от индивидов иных типологических групп [22]. Не даром же Павлов называл слабый тип трусливым. Как видно, в горниле ГБЧ каждая из трех базальных эмоций (гнев, удовольствие, страх) была последовательно размещена и закреплена, как доминирующая, за каждой конкретной типологической группой. Доминирующая эмоциональность, как известно, считается наиболее ярким показателем индивидуальных различий [12]. В особенных точках шкалы времени жизни, через строго дозированные К-периоды, у эукариотов резко изменялись жизнедеятельность и поведение. Традиционное размежевание эукариотов “человек современный” на группы по преобладающим базальным эмоциям и отличиям в поведении в тех или иных теориях, в действительности является лишь подтверждением эволюционного эха тех резких скачков в дискретном отсчете времени ГБЧ, которые зафиксированы в критических точках линейного порядка типологических групп.
Индивиды флегматоидной группы в линейном спектре “τ-типов” занимают крайнее место. Они по своим свойствам резко отличаются от индивидов предыдущих групп. Все исследователи отмечают отсутствие у флегматиков выраженного внешнего проявления эмоций. По собственной единице времени (τ>1с) флегматоиды относятся к группе “медлительных” индивидов. Недаром Павлов называл этот тип спокойным и инертным. Флегматоидная группа самая малочисленная в сравнении с другими типологическими группами. По нашим данным количество индивидов флегматоидной группы в человеческой популяции составляет около 8 % [22]. Такое же количество особей флегматоидной группы обнаружили и сотрудники Павлова среди собак [17].
Казалось бы, что на данный момент по закону дискретного отсчета времени ГБЧ, эукариоты с квантом согласования τ>1с еще не нужны, так как запрос на Cd>24 часов появится лишь в будущем. Однако, флегматоидная группа существует реально в известных пределах линейного спектра “τ-типов”, и реально существуют свободнотекущие Cd>24 часов у многих видов эукариотов животного мира при нынешнем периоде суточного вращения Земли. В созданном как бы “преждевременном” кванте согласования (τ>1с) проявилась универсальная возможность ГБЧ предвидеть будущее. Эту универсальную возможность, которая принципиально отличает живое от неживого, П.К. Анохин назвал “опережающим отражением” [2]. С учетом механизма “опережающего отражения” происхождению флегматоидной группы можно дать следующее объяснение. Создавая флегматоидную группу в линейном спектре “τ-типов”, ГБЧ как будто заложили своеобразный резерв в механизм подстройки фазы, учитывая замедление суточного вращения Земли в будущем. Так, квант времени τ=1,1с позволит эукариотам адаптироваться к периоду Cd=26,4 часа, что говорит о возможности ГБЧ вести отсчет времени от условного “нуля” в будущем еще на 576·106 лет. Но и это еще не предел хода ГБЧ, если учесть, что Cd-период у ныне существующих эукариотов может растягиваться до 30-ти часов и более [4], [18]. Убедиться в резервных возможностях ГБЧ человечество сможет лишь в далеком будущем.
Учитывая соответствующую цепочку квантов времени, с помощью которых эукариоты согласовывали свои Cd-периоды с замедлением суточного вращения Земли, в диапазоне 0,7с≤τ≤1,0с линейного спектра “τ-типов” фактически зафиксирован в спрессованном виде отрезок шкалы времени жизни, за который с момента обособления эукариотов (Т=3К), до момента появления человека современного (Т=0), в горниле ГБЧ осуществлялось последовательное усовершенствование организмов. Диапазон 1,0с<τ≤1,1с следует рассматривать как резервный отрезок спрессованной шкалы времени жизни, предусмотренный ГБЧ для нашего дальнейшего усовершенствования в будущем. Отрезок натуральной шкалы и спрессованная шкала времени жизни изображены на рисунке 1.



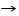 3К 2К К 0 +К
3К 2К К 0 +К
 |
0,7с 0,8с 0,9с 1,0с 1,1с
Рисунок 1: Отрезок натуральной шкалы времени жизни эукариотов и спрессованная шкала квантов согласования Cd-периодов.
В натуральном порядке типологических групп, строго локализуются “предпочитаемые” хронические неинфекционные заболевания (ХНЗ) разных органов и систем в организмах эукариотов “человек современный” [22]. Само собой разумеется, что в организме каждого индивида находятся печень, сердце, легкие, почки и другие органы. Они обеспечивают основные жизненные функции: питание, дыхание, выделение, размножение, циркуляцию веществ и перемещение [6]. Эти функции универсальны для всех эукариотов животного мира, начиная с простейших организмов (амеба), у которых уже имеет место функциональная специализация соответствующих органелл [6, с.290]. Почему же тогда один и тот же орган у индивидов определенной типологической группы оказывается наиболее уязвимым местом, а у индивидов другой группы он остается функционально надежным на протяжении всей жизни (несмотря на “усилия” современных врачей, - Б.Ц.)? Почему хронические заболевания печени присущи холерикам, сердца – сангвиникам и так далее ? [22]
Чтобы дать исчерпывающие ответы на принципиально важные для понимания психосоматического единства вопросы, нужно учесть последовательность усложнения эукариотов, зафиксированную в ряде биосферных скачков на шкале времени жизни, отсчитываемого ГБЧ. Для удобства будем оперировать отрезком шкалы времени жизни эукариотов и пропорционально спрессованной шкалой квантов согласования Cd-периодов (рисунок 1).
В организме индивидов холероидной группы наиболее уязвимым местом оказалась печень. По своим функциям печень принимает участие в пищеварении и очищении крови [25]. Кроме того, она надежно защищает организм от многих токсических веществ, которые губительно действуют на него. Вполне вероятно, что запрос именно на эту функцию возник у эукариотов еще с момента появления их в горниле ГБЧ (Т=3,3К). Согласно данным Заварзина Г.А. [10] около 2·109 лет от “нулевой” точки произошла грандиозная перестройка всей геосферно-биосферной системы на основе образования кислородной атмосферы . Тогда на Земле существовала только прокариотная биота. Появившись в точке Т=3,3К (точнее 1,9·109 лет тому), эукариоты оказались в такой геохимически опасной среде, что им на протяжении ~200·106 лет пришлось бороться за право на существование, понести множество потерь и выработать функцию защиты от токсичного окружения. Что там нынешняя экология, которой нас периодически пугают “зеленые” в сравнении с той токсичной и опасной окружающей средой? Даже не цветочки! По меткому выражению Заварзина в то время были такие экстремальные условия, в которых и “трава не растет и скот не пьет” [10, с.990]. Однако одноклеточные эукариоты выстояли и обособились в точке 3К (квант 0,7с) для дальнейшего стабильного существования, а функция противотоксичной защиты оставалась ведущей на протяжении всего периода образования холероидной группы до точки 0,8с (2К). В Венде (τ=0,87с; Т~700·106 лет) согласно данным [11], фауне в целом было присуще значительное разнообразие на уровне таксонов высокого ранга, отражающее уже практически все выбранные эволюционные направления, реализованные в «ярком» посткембрии. Согласно современным данным все эти направления были заложены намного раннее, в докембрии [10]. В конце Венда (Т~К) появляются животные с табулярным хитиновым или минеральным скелетом [11]. В момент “взрыва” скелетной революции внезапно появляются животные (это плеченогие, моллюски и членистоногие), у которых печень чрезмерно большая и характеризуется очень сложным строением [25]. «Внезапное» появление печени у животных перед началом скелетной революции очень усложняет, как подчеркивал Шмальгаузен, вопрос о ее филогении [25]. Чтобы понять загадочность филогении печени следует учесть, что ведущая функция защиты эукариотов от токсичных веществ появилась значительно раньше, а в конце Венда она лишь закрепилась за специально созданным органом – печенью. О приоритетности создания печени говорит и то, что она является самым большим внутренним органом у всех низших и высших хребетных [25]. Эволюционно созданная ведущая функция противотоксичной защиты в интервале квантов 0,7с-0,79с «отозвалась» в печени, которая остается наиболее уязвимым местом в организмах индивидов холероидной группы и поныне.
У индивидов сангвиноидной группы 0,8с ≤ τ ≤ 0,86с наиболее уязвимым органом оказалось сердце. Среди основных функций сердца (доставка с кровью веществ для питания клеток разных органов и тканей, собирание экскретов и доставка в органы выделения), наиболее существенной для понимания происхождения “предпочитаемого” ХНЗ в данной группе, является функция доставки кислорода во все клетки организма. Обратим внимание, что в точке 2К на шкале времени жизни, эукариоты, выстояв в геотоксичном окружении, резко переходят от ферментации к дыханию кислородом (квант согласования 0,8с). Чтобы кислород доходил до всех участков организма эукариотов, создается “живая” функция пульсирующей жизнедеятельности, а период пульсирования полностью соответствует кванту согласования (0,8с) с суточным вращением Земли на момент 2К. “Эхо” первобытного согласования сохранилось и поныне. Период пульса у среднестатистического индивида Tn=0,8с, а коэффициент корреляции между собственной единицей времени и индивидуальными частотами сердечных сокращений по всему спектру «τ-типов» составляет r=+0,99! [22]. Комментарии, как говорится, излишни. Изучение филогении сердца [8], [25] показывает, что оно обособляется у многоклеточных эукариотов (в сравнении с печенью) позже. Так, в периоде (конец Венда – начало Кембрия) членистоногие (Branchiata) и моллюски (Gastropoda) уже имеют обособленное сердце с предсердиями и желудочками, что является признаком древнего происхождения [5, с.490]. Сколько эукариотов с функцией пульсирующей жизнедеятельности разрывалось от перегрузок в интервале квантов согласования сангвиноидной группы, мы не выясним никогда. Но жизненно необходимая функция пульсирования предвосхитила создание пульсирующего органа – сердца. “Эхо” этого тяжелого пути до сих пор “звучит” в месте наименьшего сопротивления индивидов сангвиноидной группы. Именно в этой группе частота разрывов (в буквальном смысле) сердец наиболее высокая [21].
Для индивидов «равновесного» типа ( τ=0,9с) наиболее уязвимым местом оказались почки. Основная функция почек состоит в удалении из организма конечных продуктов обмена веществ (растворенные в воде соли и азотистые соединения). Наиболее распространенным заболеванием почек считается затруднение экскреции конечных продуктов вследствие образования в них конкрементов (преимущественно камней), и называется почечно-каменной болезнью.
Филогения почек чрезвычайно интересна. У всех эукариотов докембрия, которые существовали в Венде (кишечнополостные, беспанцирные трилобиты, “мягкие кораллы”, членистоногие) [11], имеет место диффузное удаление экскретов из организма [5]. Согласно Шмальгаузену [25], первичная почка (или туловищная почка), как обособленный орган, имеет место у хрящевых рыб и рыб с костным скелетом, которые появляются сразу же после “взрыва” скелетной революции в точке К (квант согласования 0,9). У рептилий и птиц от первичной обосабливается вторичная почка, которая уже лежит в области таза. Постоянная почка появляется лишь у млекопитающих. Таким образом, жизненно важная функция удаления экскретов из организма передается специально созданному органу - почке, филогения которой легко прослеживается у всех хребетных (предпочка, первичная и вторичная почка), имеющих минеральный скелет [25].
Как известно, основным строительным материалом для костей скелета является кальций. Принимая во внимание то, что до точки К у докембрийских животных имеет место диффузное выделение, а в точке К взрывообразно начинается скелетизация животных, можно предположить, что почка была создана как орган задержки кальция в организме, так необходимого для построения скелета. Не лишним будет напомнить, что конкременты, которые образуются в почках и стают причиной почечнокаменной болезни, по химическому составу в большинстве – оксалаты кальция. “Эхо” этой “тайной” функции почек сохранилось у индивидов “равновесного” типа и зафиксировано квантом согласования 0,9с на спрессованной шкале времени жизни (рисунок 1). “Дефицитный” материал для построения минерального скелета задерживается и сейчас в организмах “равновесных” и накапливается в наиболее ответственном органе – почке, в виде соединений кальция.
Меланхолики, согласно Гиппократу, находятся в чрезвычайно невыгодном положении, так как они «болеют» всеми «предпочитаемыми» ХНЗ, которые свойственны холерикам, сангвиникам, «равновесным» и, даже, флегматикам (болеют по типу флегматиков, - по Гиппократу). В этом нет ничего удивительного, так как все органы основных функций жизнедеятельности организма на момент появления кроманьонца были уже окончательно сформированы. Врачам тип «больного» меланхолика достаточно хорошо известен. Благодаря внутренней высокой чувствительности его состояние отображается в синдроме «все болит». И действительно, по объективным показателям в определенном возрасте у индивидов этого типа имеют место функциональные расстройства печени, почек, сердца, желудка и кишечника (стрессогенные язвы, о которых рассказал всему миру Г. Селье), повышение артериального давления и так далее. Им же свойственны и нервно-психические расстройства истероидного и шизоидного типа [22].
Но наиболее уязвимой у индивидов меланхолоидной группы оказалась сердечно-сосудистая система, точнее сосуды головного мозга, которые разрываются от резкого скачка кровяного давления. Напомним, что “insult” в переводе с латыни – скачек. Что же касается инфарктов, то у индивидов меланхолоидной группы они в большинстве микро- и мелкоочаговые. Меланхолоиды (0,96с ≤ τ ≤ 1,0с) могут перенести последовательно ряд таких инфарктов без особых угроз для жизни [21]. Инсульт, наоборот, может быть фатальным. Почему именно так распорядилась природа по отношению к меланхоликам? Для объяснения, прежде всего отметим, что П. Лесгафт в свое время разработал оригинальное толкование природы индивидуальных различий, которое не было воспринято должным образом. Он считал, что в основу темперамента заложены свойства системы кровообращения, и отдельно толщина стенок (подчеркнуто нами, - Б.Ц.) кровеносных сосудов и диаметр их просвета [14,с.105].
Наиболее уязвимые инсультогенные типы находятся в диапазоне 0,96с≤τ≤1,0с спрессованной шкалы квантов согласования (смотри рисунок 1). Этому диапазону на шкале времени жизни соответствует период 1/2К–0. Именно в нем по палеонтологическим данным на промежутке 1/2K–1/4K появляются самые первые представители теплокровных – аллотерии, а в момент их исчезновения (~1/8K) обосабливается ветвь приматов. Завершающим звеном в последовательной цепи эволюционных скачков на этой ветви стал вид “человек современный” с наиболее развитым головным мозгом. Не трудно понять, что в горниле ГБЧ после загадочного большого мезозойского вымирания в окрестности точки Т12=1/8K , когда внезапно исчезли динозавры [23], именно в ветви приматов началось интенсивное усовершенствование центральной нервной системы в направлении увеличения массы головного мозга. Фактически начался процесс усовершенствования центрального механизма часов. В свою очередь, разрастание головного мозга требовало более интенсивного питания его кровью за счет повышения кровяного давления и уменьшения толщины стенок кровеносных сосудов. “Эхо” от этих “проб и ошибок” отозвалось в тонких сосудах, высоком артериальном давлении и тахикардии у индивидов меланхолоидной группы [21].
У представителей флегматоидной группы наиболее уязвимой оказалась система пищеварения и, особенно, желудок. Несмотря на наидревнейшее происхождение (скорее всего, одновременно с функцией противотоксичной защиты), система пищеварения – питания флегматоидов своей уязвимостью возможно предупреждает (как Минздрав) о насущных усложнениях, которые возникнут в будущем? Но этот вопрос на данное время остается открытым.
Чтобы картина создания «человека современного» в горниле ГБЧ была полной, необходимо рассмотреть вопрос о происхождении нашего наиболее мощного, по сравнению с другими млекопитающими, интеллектуального потенциала. В традиционном толковании о роли труда в процессе превращения обезьяны в человека имеется достаточно много известных противоречий и расхождений [24]. С нашей точки зрения они возникли в результате существенного пробела, обусловленного тремя качественными скачками в переходе от вида «человек разумный» к виду «человек современный» [23]. Подобные скачки в ветви приматов после большого мезозойского вымирания существенно отличаются от скачков в эукариотной биоте до точки 1/8K. До этой точки, как показано в нашем отдельном исследовании [23], продолжительность существования различных представителей фауны определялась их видовой возможностью к подстройке Cd-периода соответственно замедлению суточного вращения Земли. Так, например, выбрав резерв позволенного затягивания Cd-периода, исчезли трилобиты, ракоскорпионы, кистеперые рыбы, брахиоподы и вся цепочка “стегоцефалы – котилозавры – текодонты – динозавры”. С ветви приматов резко началось усовершенствование центральной нервной системы и, особенно, головного мозга. Этот орган, как разумеется, стал наиглавнейшим часовым механизмом у приматов, что позволяло им подстраивать свои Cd-периоды к замедлению суточного вращения Земли с нарастающей точностью. Поэтому общая ветвь приматов сохраняется в цепи дискретных переходов хода ГБЧ. Исчезает одна его разновидность, но сразу же появляется другая, более совершенная разновидность, способная синхронизировать фазы собственных околосуточных ритмов в соответствии с удлинением смены световых и темновых периодов суток с все более нарастающей точностью. Уровень нашего видового интеллектуального потенциала, взлелеянный в горниле ГБЧ, строго определяется не только точностью подстройки фазы индивидуального Cd-периода, но и качеством хода индивидуальных часов личности [22]. В отдельном исследовании [7] было строго доказано, что показатель качества хода индивидуальных часов (Q) высоко коррелирует (r = – 0,97) с уровнем психометрического интеллекта и определяет врожденный интеллектуальный потенциал субъекта, который традиционно в психологии называется “загадочным фактором g”.
В заключение отметим следующее: эукариоты «человек современный» со всем разнообразием индивидуальных свойств психики созданы и выстроены в натуральный порядок четырех типологических групп (подчеркнуто нами, – Б.Ц.) в горниле ГБЧ по закону дискретного отсчета времени существования живого вещества. Вся совокупность проб и ошибок поиска вектора усовершенствования нашего психосоматического единства последовательно отчеканилась в линейном спектре “τ-типов”. Сам спектр в пределах 0,7с≤τ≤1,1с оказался компактно спрессованным отрезком времени существования эукариотной биоты с соответствующим резервом нашего дальнейшего усовершенствования в будущем. Закон дискретного отсчета времени ГБЧ позволяет говорить о реальности появления нового (или новых) вида “Homo futurus”, способного даже к искусственному самоусовершенствованию (подчеркнуто нами, – Б.Ц.) своего психосоматического единства. Минимальный отрезок времени, отведенный для этого ГБЧ, известен.
Человечество из поколения в поколение воспроизводит индивидов по психогенетическому закону в стойких пределах линейного спектра “τ-типов” и сохраняет натуральный порядок четырех типологических групп. В основании этого закона лежит механизм дискретных колебаний маятника репродукции от одного крайнего состояния ( τ=0,7с) к противоположному крайнему состоянию ( τ=1,1с) [20]. По психогенетическому закону каждый новорожденный индивид уже принадлежит (Б.Ц.) к определенной типологической группе и занимает в ней свое собственное место, согласно врожденному “τ-типу”, с соответственно “смешанными в надлежащем отношении частей” формально-динамическими свойствами психосоматического единства. Все, с чем приходит каждый индивид в мир своего собственного существования, уже заложено в его психосоматическом единстве. В нем действительно смешаны в соответствующую пропорцию и формально-динамические свойства, и ведущий эмоциональный фон, окрашивающий поведение, и место наименьшего сопротивления в организме, и природный умственный потенциал человека. Каждая пропорция смешивалась и создавалась в горниле ГБЧ в той последовательности, которая зафиксирована в натуральном порядке четырех типологических групп, а они оказались лишь эхом ряда постепенных и скачкообразных качественно-количественных изменений в процессе усовершенствования эукариотов.
Мы, эукариоты наивысшего уровня оганизации, сознательные индивиды, созданные в горниле ГБЧ по закону дискретного отсчета времени биосферы, оказались всего лишь его наиболее универсальными “стрелками”, способными “объективно” обосновывать различие между прошлым и будущим, опираясь на квант собственного “действительного настоящего”. Рефлексивно, на уровне сознания, мы легко фиксируем и момент своего рождения, и момент падения Трои, и момент исчезновения динозавров, и, даже, предполагаемый момент рождения Вселенной, как ряд последовательных изменений с появлением, становлением и исчезновением. Но мы остаемя беспомощными и бессильными перед вопросом: “как или почему началось время?”, которое равнозначно вопросу: “как или почему началась жизнь?”.
Изложенное выше решение задачи Фр. Гальтона было построено на учении В.И. Вернадского, согласно которому в ходе первой биогеохимической революции начало функционировать первичное живое вещество на Земле. Но, как известно, причина начала саморазвития живого вещества еще не получила исчерпывающего научного доказательства.
На этот, наверное, наисложнейший вопрос у человечества есть и другой ответ: все живое, по Библии, было создано Всевышним. Если мы и созданы Богом, то Глобальные биологические часы, бесспорно, были его творческой мастерской, а инструментом было время, щедро отведенное нам в достаточном количестве для самоосознания всей уникальности и нашего собственного бытия, и нашего собственного существования.
Литература.
1. Анастази А. Дифференциальная психология. Конспект и комментарий Б.М. Теплова / В сб.: Проблемы дифференциальной психофизиологии. Москва, 1969, т. 4 –с. 304- 339.
2. Анохин П.К. Очерки по физиологии функциональных систем. – Москва, 1975 – 476 с.
3. Ашофф Ю. Свободнотекущие ритмы / Биологические ритмы. – Москва, 1984, - т. 1, с. 15-20.
4. Биологические часы. – Москва, 1964 – 694 с.
5. Бородулин Ф.Р. Лекции по истории медицины. – Москва, 1955. - с. 14-28.
6. Вилли К., Детье В. Биология. – Москва, 1974. – 822с.
7. Головина Н.В., Цуканов Б.И. Прирожденная мера интеллекта // Вестник Одесского государственного университета, 1999, т.4, вып. 2, с. 87-91.
8. Догель В.А. Сравнительная анатомия беспозвоночных. – Ленинград, 1939. – 324 с .
9. Друщиц В.В., Обручева О.П. Палеонтология. – Москва, 1962. – 379с.
10. Заварзин Г.А. Становление биосферы // Вестник РАН. – 2001. – т. 71. - №11. – с.988-997.
11. Мороз С.А. История биосферы Земли. Книга 2, Киев, 1996, 365с. (на украинском языке).
12. Небылицын В.Д. Психофизиологические исследования индивидуальных различий. – Москва, 1976. – 336 с.
13. Павлов И.П. Полное собрание сочинений. – Москва; Ленинград, 1951. – Т.3 – Кн. 1. – с. 381-420.
14. Психология. Под ред. проф. Трофимова Ю.Л. - Киев, 1999, с. 104-105.
15. Равич-Щербо И.В. и др. Психогенетика. – Москва, 1999. – 447 с.
16. Симонов П.В. Естественнонаучные основы индивидуальности // Журнал высшей нервной деятельности. – 1981. – Т. 31. - №1. – с. 12-24.
17. Теплов Б.М. Проблемы индивидуальных различий. – Москва, 1961. – с. 347-495.
18. Уинфри А. Время по биологическим часам. – Москва, 1990. – 208 с.
19. Цуканов Б.И. Фактор времени и природа темперамента // Вопросы психологии. – 1988. - №4. – с. 129-136.
20. Цуканов Б.И. Психогенетические закономерности воспроизводства индивидов в типологических группах // Вопросы психологии. – 1989. - №4. – с. 149-153.
21. Цуканов Б.И. Фактор времени и проблема сердечно-сосудистых заболеваний // Психологический журнал. – 1989. – Т.10. - №1. – с. 83-88.
22. Цуканов Б.И. Время в психике человека. – Одесса, 2000. – 220 с.
23. Цуканов Б.И. Глобальные биологические часы эволюции // Правовое государство, 2000, №2, с. 81-89 (на украинском языке).
24. Чайковский Ю.В. Элементы эволюционной диатропики. – Москва, 1990. – 270 с.
25. Шмальгаузен И.И. Основы сравнительной анатомии позвоночных животных. – Москва, 1947. – 520 с.