
Подготовил Студент Факультета ФЕНМиТ Манченко Дмитрий Группа ПОБХ (б) 61








Реферат
по теме: «Эволюция копытных млекопитающих.»
Подготовил Студент
Факультета ФЕНМиТ
Манченко Дмитрий
Группа ПОБХ (б) 61
Эволюция копытных млекопитающих
Содержание
1. Введение
2. Эволюция непарнокопытных млекопитающих на примере вида Equus.\
3. Эволюция парнокопытных млекопитающих.
4. Заключение
Введение.
Копытные млекопитающие дают очень обильный и разнообразный палеонтологический материал. Все принадлежавшие сюда животные были растительноядны и за редкими исключениями (Anoplotherium, бегемоты) вели наземную жизнь. Генетически самые древние копытные (Protungulata) стояли весьма близко к креодонтным хищникам. Череп Protungulata напоминал таковой у креодонтов и был снабжен продольным гребнем; зубная система состояла из 44 гетеродонтных зубов. Верхние коренные по типу были трехбугорчатые, как у древних хищников; нижние коренные—туберкул о-секториальные. Самую древнюю группу Protungulata составляют нижнеэоценовые североамериканские Mioclaenidae, от которых, к сожалению, сохранились лишь остатки черепов и челюстей с зубами. Лучше других первичнокопытных известен североамериканский эоценовый ф е н а к о д . Это небольшое животное, ростом с теленка, отличалось примитивным строением пятипалых конечностей. Относительно древности и неизмененности структуры передней ступни фенакода писал О. Абель (1914), который сравнивает эту ступню с кистью руки человека, говоря, что у людей эта кисть устроена наиболее примитивно в ряду всех современных млекопитающих. Морда фенакода была вытянута в длинный хоботок; хвост у этого животного был сильно удлинен; зубы по структуре напоминали креодонтов.
Первичнокопытные, или кондилартры, послужили основным стволом, из которого в дальнейшем развилось богатое разнообразие остальных Ungulata. Из них вышли современные низкоорганизованные даманы, сохранившие многие примитивные черты (например, в структуре конечностей). Особую угасшую ветвь составляет нижнеолигоценовый отряд Embrithopoda. Огромный, с крупного массивного носорога Arsinoitherium zitteli был обнаружен в слоях Фаюма в Египте. Один ствол приводит к хоботным, другие—к парно- и непарнокопытным. Целую группу отрядов (Typotheria, Toxodontia, Entelonychia, Astrapotherioidea) составляют давно угасшие и вымершие южноамериканские копытные, объединяемые в широкий отдел Notungulata. Также совершенно вымерли миоценовые европейские и североамериканские непарнокопытные Ancylopoda, эоценовые североамериканские Amblypoda и Litopterna (из эоцена Южной Америки).
Рассмотрим несколько наиболее ярких примеров эволюционного процесса в ряду некоторых групп копытных. Очень наглядную картину эволюции дают бугорчатозубые мастодонты.
Рис.1
Бугорчатозубые мастодонт.

Самой древней формой среди них считается по справедливости меритерий (Moeritherium lyonsi), обнаруженный в среднем эоцене близ Фаюма (в Египте). Это животное величиной с тапира было лишено хобота, обладало четырехбугорчатыми зубами и удлиненной структурой резцов. Из верхнего эоцена Фаюма сохранился более крупный Palaeomastodon с вытянутой нижней челюстью. Дальнейшими представителями эволюционного ряда можно назвать Bunolophodon angustidens из миоцена Европы и Северной Америки В. longirostris и В. arvernensis (миоплиоцен Европы).
Относительно полно изучена история настоящих слонов. Зубы азиатского слона (Elephae maximus) по форме многочисленных пластинок стоят ближе всего к индийскому плиоценовому Е. hisudricus. Мамонт плейстоценовые находки которого многочисленны в Евразии, отличался наиболее специализированными и сложными по структуре зубами среди прочих хоботных. Интересно, что в вечно мерзлой почве севера Сибири установлены находки мамонтов, сохранившихся с мясом, шерстью и внутренностями. Животные, по-видимому, проваливались иногда между льдами в болотистом грунте тундры и замерзали, сохраняясь так в течение многих тысячелетий. Изучение мамонта дает ценнейший материал для установления анатомии и экологии этих вымерших гигантов. Африканский слон произошел от других предков. По структуре зубов он стоит наиболее близко к вымершему роду Stegodon (из сиваликских отложений близ Пикерми в Греции).
Эволюция копытных млекопитающих на примере вида Equus.
Одну из наиболее ярких страниц в палеонтологической летописи и чрезвычайно показательный пример эволюционного процесса представляет история развития лошади. Палеонтологические остатки свидетельствуют с полной отчетливостью, как от общего типа пятипалой конечности древних копытных могла возникнуть однопалая конечность современных лошадей.
Equus, или лошадь обыкновенная – современный и самый распространенный вид копытных на Земле. Благодаря большому количеству ископаемых останков и богатой палеонтологической истории, на примере этого вида можно рассмотреть особенности и причины эволюции копытных млекопитающих.
Современной лошади предшествовали ещё 7 видов, существовавших в разные периоды. Сравнивая различия между ними и выясняя, почему данные изменения были приобретены, можно понять особенности эволюции копытных млекопитающих.
Самым древним предком современной лошади является Эогипус (Eohippus).
Рис.2 Эогуппус (Eohippus)

Эогиппусы обитали на планете около 50 миллионов лет назад. Эти животные по внешнему виду напоминали лошадь, но ростом они были гораздо меньше – примерно как современные кошки. Благодаря сходству с лошадьми эогиппусы получили свое название, так как «эос» означает «заря», а «гиппос» переводится как «лошадь». Высота эогиппусов в холке не превышала 50 сантиметров, но самые маленькие особи достигали лишь 25 сантиметров. Эти животные могли быстро бегать, поскольку у них были длинные и хорошо развитые ноги. Они жили в болотистой местности, а удерживаться на тонкой поверхности им помогали широко расставленные пальцы. Между пальцами была мягкая подушка, как на лапе современной собаки. Благодаря такому строение стопы эти лошади легко передвигались по болотистой местности. Челюсти были снабжены 44 крепкими зубами, с помощью которых эогиппусы легко пережевывали жесткую растительность. Глаза у этих животных сидели довольно близко, поэтому боковое зрение отсутствовало. Тело эогиппуса покрывала жесткая короткая шерсть. Окрас был полосатым или пятнистым. Он выполнял роль камуфляжа, животные могли скрываться в траве от хищников. На основании изучения останков, был сделан вывод, что эти доисторические лошади обитали в тропических лесах. Данное предположение подтвердилось благодаря обнаруженным рядом с останками эогиппусов костей обезьян.
Далее идет промежуточный вид Орогиппус (Orohippus), но я не вижу смысла заострять на нём внимание, так как различия между Эогипусом малы.
Рис.3 Орогиппус (Orohippus) и Мезогиппус (Mesohippus).

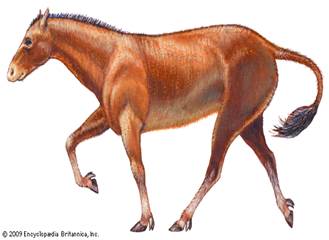
Перейдем сразу к третьему по древности предку – Мезогиппусу (Mesohippus). Мезогиппусы, как и все лошади, были травоядными животными, жили в стадах. Они были приспособлены к жизни на открытых просторах, однако большую часть времени проводили и в лесах. Передвигались рысью, что свойственно и современным лошадям. Мезогиппусы были значительно меньше современных лошадей, они были размером с волка. Они не имели копыт: на каждой конечности было по три пальца, причём центральный был больше остальных. Коренные зубы Мезогиппусов были лишены цемента.
Здесь уже можно провести некоторые сравнения: например, размер. Как мы помним, Эогиппус был размерами примерно с кошку (50 см), а Мезогиппус явно обогнал своего предка, вымахав до размера волка (80-100 см). Следующее отличие – конечности. Эогиппус имел пять пальцев на ногах, что позволяло ему без проблем передвигаться по трясине и болотистой местности, Мезогиппус же имел уже три пальца на ноге, причем центральный был больше, что является переходным признаком от кистехождения к пальцехождению. Доподлинно не известно, сохранил ли Мезогиппус покровительственный окрас своего предшественника, но можно с уверенностью сказать, что это животное все ещё не могло передвигаться достаточно быстро, чтобы скрыться от хищников на открытом пространстве.
Следом идет ещё один промежуточный вид, не имеющих выделяющихся отличий от предыдущего, зато его потомок, Парагиппус (Parahippus)
Рис.4 Парагиппус (Parahippus)
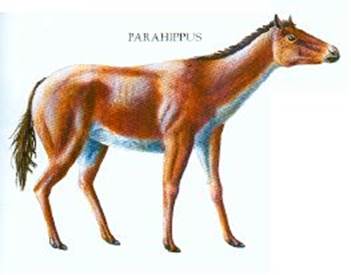
представляет для нас интерес. Это животное имело рост чуть больше метра в холке. Основными отличиями были удлиненные ноги с подобием копыт, и вытянутая морда.
Заострим внимание на конечностях: средний палец, который для животного являлся опорным, был покрыт роговым наростом, что уже можно назвать подобием копыта. Такие изменения связаны с изменением климата: джунглей, лесов и болот стало меньше, что вынудило Парагиппусов выйти на прерии, где покровительственный окрас уже не спал. В такой ситуации животным нужно было увеличить скорость передвижения, о чем говорят удлиненные ноги. Морда же стала длиннее в виду изменения дыхательной системы – при быстром беге телу требуется куда больше кислорода. В следствие этого, носовые отверстия увеличились, чтобы за один вдох получать больше кислорода.
Следующие потомки современной лошади, на которых мы заострим внимание – Merychippus и Pliohippus.
Рис. 5 Меригиппус (Merychippus) и Плиогиппус (Pliohippus) 

Они уже мало чем отличались от современных представителей вида. Эти животные имели по одному ороговевшему пальцу на ногах и, по сути, являлись первыми полностью копытными представителями вида Equus.
Переход на прерии не прошел даром – лошади приобрели копыта, увеличили размер тела, покровительственная окраска пропала почти полностью, зубы приспособились к пережевыванию твердой и сухой пищи, глаза были посажены низко и имели обзор почти 340 градусов. Дыхательная и сердечно-сосудистая системы тоже претерпели изменения от перехода к жизни в прериях – легкие увеличились, а сердце стало более сильным, что позволяло животным как передвигаться быстро на короткие дистанции, так и совершать длительные миграции.
Эволюция парнокопытных млекопитающих.
Параллельно с непарнокопытными, коими и являются лошади, шла эволюция парнокопытных представителей млекопитающих.
Для начала хотелось бы отметить некоторые общие особенности, присущие парнокопытным млекопитающим.
Характеристика отряда парнокопытные: Копытные с числом пальцев, как на передних, так и на задних конечностях, два или четыре. Сильнее других развиты третий и четвертый пальцы. Между ними проходит ось симметрии конечности, и эти два пальца несут на себе основную тяжесть тела животного. Второй и пятый пальцы в той или иной степени недоразвиты, иногда совсем отсутствуют. Гомологичные фаланги функционирующих пальцев имеют зеркальное сходство (как бы отражение одна другой). Третья фаланга сжата с боков и имеет несимметричную треугольную форму. Третий вертел (trochanter tertius) на бедре отсутствует. Шейка бедра у парнокопытных ясно отделяет суставную головку от тела кости. Межвертельный гребень идет от большого к малому вертелу и ограничивает с латеральной и дистальной стороны вертельную ямку. Таранная кость имеет два суставные блока: проксимальный для сочленения с костями голени и дистальный. Пяточная кость парнокопытных, помимо таранной, всегда сочленяется также с малой берцовой или ее рудиментом.