
Модель строения биологических мембран Даниелли–Девисона








1 – углеводородные гидрофобные цепочки; 2 – полярные
гидрофильные группы молекулы; 3 – полярные поры, по которым
вещества диффундируют в клетку
В 1959 г. Робертсон Дж. Д. предположил, что все клеточные мембраны построены по единому принципу, и высказал концепцию унитарной (или единообразной) мембраны

модель во многом сходна с классической моделью Даниелли Дж.: основу мембраны составляет липидный бислой, а нелипидные компоненты (прежде всего белки) в полностью развернутой конформации лежат на поверхности бислоя, связываясь с липидами за счет электростатических и гидрофобных взаимодействий. В модели Робертсона нашла отражение еще одна важная структурная особенность мембраны – ее асимметрия. Согласно его теории БМ содержит белки, которые связанны электростатически; на наружной поверхности БМ находятся гликолипиды.
В 1972 г. Сингер и Николсон предложили жидкостно-мозаичную модель мембраны, согласно которой белковые молекулы плавают в жидком фосфолипидном бислое. Они образуют в нем как бы своеобразную мозаику, но поскольку бислой этот жидкий, то и сам мозаичный узор не жестко фиксирован; белки могут менять в нем свое положение.
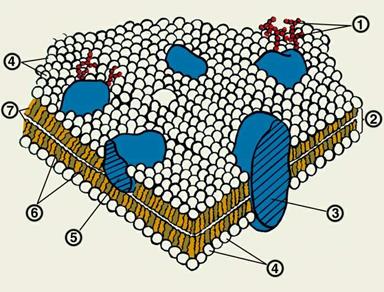
1 – углеводные фрагменты гликопротеидов; 2 – липидный бислой;
3 – интегральный белок; 4 – «головки» фосфолипидов;
5 – периферический белок; 6 – холестерин;
7 – жирнокислотные «хвосты» фосфолипидов
Согласно жидкостно-мозаичной модели:
1) Структурной основой биомембран является липидный бислой, в котором углеводородные цепи молекул фосфолипидов находятся в жидкокристаллическом состоянии.
2) В липидный бислой, имеющий вязкость растительного масла, погружены или встроены молекулы белков, способные передвигаться по мембране.
3) Толщина мембран составляет около 7 нм.
4) Гидрофильные головы фосфолипидных молекул обращены наружу — в сторону водного содержимого клетки и в сторону наружной водной среды. Гидрофобные хвосты обращены внутрь — они образуют гидрофобную внутреннюю часть бислоя.
5) Белки сохраняют связь с мембраной, поскольку в них есть участки, состоящие из гидрофобных аминокислот, взаимодействующих с гидрофобными хвостами фосфо-липидов; вода из этих мест выталкивается. Другие участки белков гидрофильны. Они обращены либо к окружению клетки, либо к ее содержимому, т. е. к водной среде.
6) Некоторые мембранные белки лишь частично погружены в фосфолипидный бислой, тогда как другие пронизывают его насквозь.
7) К некоторым белкам и липидам присоединены разветвленные олигосахаридные цепочки, играющие роль антенн. Такие соединения называются соответственно гликопротеинами и гликолипидами.
8) В мембранах содержится также холестерол. Подобно ненасыщенным жирным кислотам он нарушает плотную упаковку фосфолипидов и делает их более жидкими. Это важно для организмов, живущих в холодной среде, где мембраны могли бы затвердевать. Холестерол делает мембраны также более гибкими и вместе с тем более прочными. Без него они бы легко разрывались.
5. Химический состав биологических мембран: соотношение белков и липи-дов. Классификация мембранных белков.
Мембрана не статическое образование, а благодаря жидкокристаллической структуре она является двухслойным раствором, в котором липиды способны диффундировать как параллельно поверхности мембраны, так и из одного монослоя в другой.
В структуру мембран обязательно входят белки и их состав варьирует в зависимости от функции той или иной мембраны.
В зависимости от прочности связи с мембраной различают:
· периферические,
· интегральные.
Интегральные белки располагаются между липидами монослоя или пронизывают весь бислой, часто возвышаясь над поверхностью мембраны. Периферические белки связаны с мембранами электростатическими и водородными связями и часто взаимодействуют таким образом с интегральными белками.
Белки выполняют следующие функции:
· транспорт (трансмембранный перенос веществ);
· преобразование энергии (ферменты дыхательной цепи);
· коммуникативную (рецепторные белки связывают клетку с окружающей средой);
· ряд специфических функций.
Углеводы в биологических мембранах находятся в соединении с белками (гликопротеины) и липидами(гликолипиды). Углеводные цепи белков представляют собой олиго- или полисахаридные структуры, в состав которых входят глюкоза, галактоза, нейраминовая кислота, фукоза и манноза. Углеводные компоненты биологической мембраны открываются в основном во внеклеточную среду, образуя на поверхности клеточных мембран множество ветвистых образований, являющихся фрагментами гликолипидов или гликопротеидов. Их функции связаны с контролем за межклеточным взаимодействием, поддержанием иммунного статуса клетки, обеспечением стабильности белковых молекул в мембране. Многие рецепторные белки содержат углеводные компоненты.
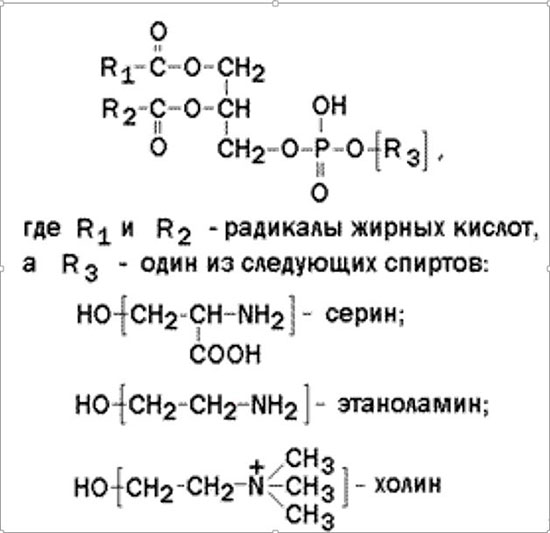
Фосфолипиды — один из основных компонентов плазматической мембраны эукариот и бактерий. Являются сложными эфирами жирных кислот («хвосты») и многоатомных спиртов (преимущественно глицерола, сфингозина и инозитола), соединенных через остаток фосфорной кислоты с добавочной группой («головкой»). Глицерофосфолипидов в мембранах по массе больше всего, и это наиболее пластичный и текучий их компонент. В зависимости от типа головки эти липиды делят на классы:
· фосфатидилхолины (головка — холин),
· фосфатидилглицеролы,
· фосфатидилэтаноламины и т.д.
Полное название липида включает также наименование жирных кислот, образующих хвосты, например, олеиновая и пальмитиновая кислоты образуют пальмитоилолеилфосфатидилхолин (ПОФХ), а две молекулы пальмитиновой кислоты — дипальмитоил фосфатидилхолин (ДПФХ).
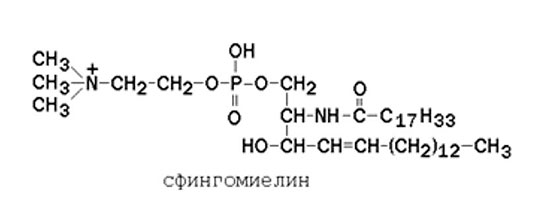
Сфингофосфолипиды также имеют два ацильных хвоста, но только один из них принадлежит жирной кислоте: второй относится к сфингозину. Один из таких липидов — сфингомиелин (в обилии встречается в миелиновой оболочке аксонов) — является важным компонентом клеточной мембраны, придавая ей жесткость и уникальные физико-химические свойства. Самые простые сфингофосфолипиды называются церамидами.
Холестерол (он же холестерин) — другой важный липид клеточной мембраны, стабилизирующий ее текучесть, — является отличительной чертой мембран эукариот (его нет ни у бактерий, ни у архей). Кроме стабилизации мембран холестерол является предшественником половых гормонов и витамина D.
Гликолипиды, имеющие в качестве полярной головки один или несколько остатков сахаров, также являются компонентами мембран (преимущественно внешнего монослоя). Основной их формой являются гликосфинголипиды, благодаря чему эти липиды, как правило, колокализованы со сфинголипидами (а именно, расположены в жидкой упорядоченной фазе). Основная их роль — межклеточное распознавание, и кроме пользы (например, определения групп крови) эта их функция может быть и вредна: многие ганглиозиды являются рецепторами бактериальных токсинов и вирусов.
К мембранным белкам относятся белки, которые встроены в клеточную мембрану или мембрану клеточной органеллы или ассоциированы с таковой. Около 25 % всех белков являются мембранными.[1]
Биохимическая классификация
По биохимической классификации мембранные белки делятся наинтегральные и периферические.
· Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентовили неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими.
· Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором (например, с низким или высоким pH, с высокой концентрацией соли или под действием хаотропного агента). Эта диссоциация не требует разрушения мембраны.
Мембранные белки могут быть встроены в мембрану за счёт жирнокислотных или пренильных остатков либогликозилфосфатидилинозитола, присоединённых к белку в процессе их посттрансляционной модификации.
6. Строение основных липидов биомембран.
Компонентами липидов являются остатки жирных кислот и одно- или многоатомных спиртов. Примеры жирных кислот, встречающихся в составе липидов мембран.

Основные особенности строения жирных кислот, входящих в состав природных жиров:
· они содержат чётное число атомов углерода (С16 – С20);
· двойная связь в ненасыщенных жирных кислотах располагается между 9 и 10 атомами углерода;
· в полиненасыщенных жирных кислотах двойные связи разделены метиленовыми группами (СН=CH-CH2-CH=CH), то есть являются несопряжёнными;
· двойные связи находятся в цис-конформации, что приводит к изгибу углеводородной цепи.
Большинство липидов в мембранах млекопитающих представлены:
· фосфолипидами,
· гликосфинголипидами,
· холестеролом.
Фосфолипиды в составе мембран подразделяются на две группы: глицерофосфолипиды и сфингомиелины.
Глицерофосфолипиды – представляют собой сложные эфиры трёхатомного спирта глицерола, двух остатков жирных кислот и фосфорилированного аминоспирта. Общая формула глицерофосфолипида
Наиболее распространённым глицерофосфолипидом мембран является фосфатидилхолин:
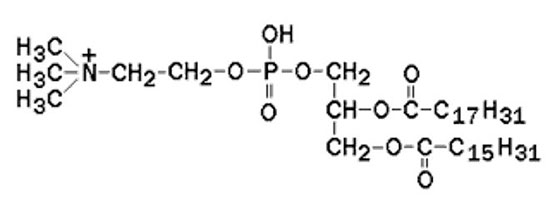
В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой). Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина. Соединение сфингозина и жирной кислоты получило название церамид.
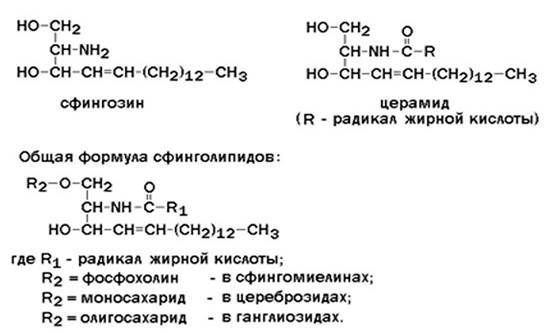
В сфингомиелинах водород гидроксильной группы у первого углеродного атома в церамиде замещён на фосфохолин. Пример сфингомиелина, содержащего остаток олеиновой кислоты:
Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы:
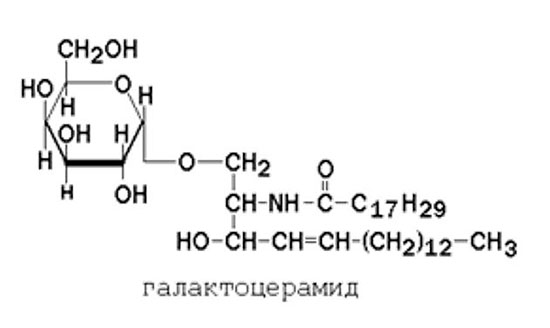
а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота.
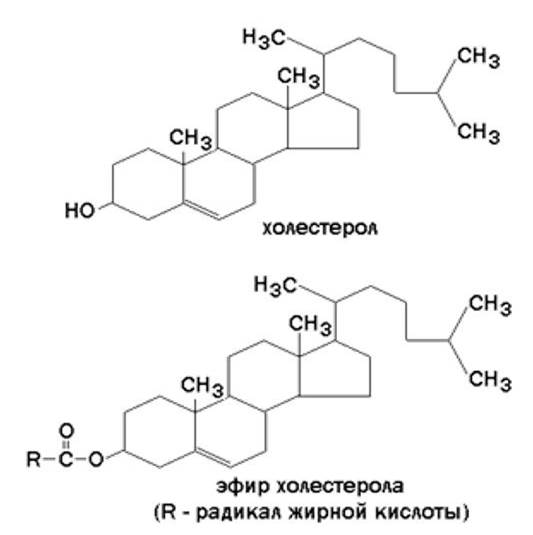
Холестерол – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани.
Характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина.
Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна.
Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду.
Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран.
9. Искусственные фосфолипидные мембраны как модели биологических мембран (липосомы, протеолипосомы).
Искусственные липидные мембраны, имеющие двуслойное строение, оказались во многих отношениях сходными с биологическими мембранами. Искусственные мембраны получаются при контакте смеси фосфолипидов и нейтральных липидов, растворенных в органических растворителях, с водой. Бимолекулярные липидные мембраны (БЛМ), называемые также бислойными или черными липидными мембранами, представляют собой широко используемую экспериментальную модель, которая позволяет воспроизводить в искусственных условиях многие свойства и характеристики биологических мембран. Как и биологические мембраны, они представляют собой замкнутые системы, что делает их пригодными для изучения пассивного транспорта ионов и малых молекул через липидный бислой.
Липосомы — сферические везикулы, имеющие один или несколько липидных бислоев. Образуются в смесях фосфолипидов с водой. Внутри липосом содержится вода или раствор, в котором проводилась ультразвуковая обработка. В отличие от БЛМ, липосомы достаточно стабильны и не содержат органических растворителей. Состав липидов в липосомах можно произвольно варьировать и таким образом направленно изменять свойства мембраны. Благодаря возможности реконструкции мембраны из ее основных компонентов удается моделировать ферментативные транспортные и рецепторные функции клеточных мембран. В липосомы можно ввести антигены, а также ковалентно присоединить антитела и использовать их в иммунологических исследованиях. Они представляют собой удобную модель для изучения действия многих лекарственных веществ, витаминов, гормонов, антибиотиков и т. д.
В настоящее время хорошо разработаны методы включения функционально-активных мембранных белков в липосомы. Такие искусственные белково-липидные структуры обычно называются протеолипосомами.
Эффективность встраивания большинства белков компонентов в искусственные мембранные системы резко зависит от липидного состава мембран, pH, солевого состава, температуры и т. д. Система протеолипосомы — коллодиевая пленка , первоначально разработанная для изучения бактериородопсина, была затем использована при исследовании целого ряда других мембранных преобразователей энергии.
Существует два основных типа искусственных мембран: классические плоские, сферические мембраны различного размера. Для получения искусственных мембран используют: различные фосфатиды, нейтральные глицериды, смеси липидов биологического происхождения, добавляя к ним холестерин, а-токоферол и другие минорные добавки.
Потенциальная ценность искусственных мембран для исследований зависит от возможности включения в них природных белков, в особенности тех, которые обладают транспортными свойствами. Липосомы, состоящие из белков и липидов, стали получать в 60-е гг. Термин протеолипосомы был введен В. П. Скулачевым. В настоящее время разработан целый ряд методов приготовления различных типов липосом и протеолипосом, а также их стандартизации по размерам, структуре, гомогенности, стабильности и другим характеристикам. Липосомы используют для доставки в клетку лекарственных и химических соединений, стабилизации ферментов в инженерной энзимологии, введения в клеточные мембраны молекул зондов, модифицирующих и моделирующих их поверхность. Большой интерес для генной инженерии и медицины представляют работы по введению в клетки при помощи липосом нуклеиновых кислот и вирусов.
Из книги-Ревин: Важную роль в изучении биол мембран играет моделирование-формирование искусственных моно- или бислойных липидных мембран и протеолипосом базирующееся на том, что амфипатические молекулы липидов способны образовывать на границе раздела вода-органический растворитель мономолекулярные пленки. В настоящее время широко исп-ся методы получения моно- и бислойных липидных мембран, а также методы формирования замкнутых пузырьков-везикул,липосом. В такие мембраны вводят белки или мелкеи фрагменты мембран, выделенных из клетки-протеолипосомы. Так как липидный бислой-обязательный компонент биол мембраны, результаты, полученные при исследовании физических характеристик модели мембраны и ее проводимости, имеют большое значение для понимания ряда процессов организации и функционирования нативных биол мембран. Возможности такого рода моделирования еще более расширяются при формировании протеолипосом.
10. Состояние воды в клетке. Свободная и структурированная вода в клетке
С водой связаны многие структурно-функциональные свойства мембран, а также процессы стабилизации и формирования мембран. Молекула воды- диполь, т е на одной стороне -, на другой +. Благодаря такому св-ву молекула воды в клетке образует динамическую структуру.
Ф-ии воды:-расворитель орг и неоргн в-в
-дисперсионная система
-метаболизм
-терморегуляция
-тургор
-участвует в стабилизации макромолекул
-растворитель в клетке
Вода входит в состав мембран и делится на: свободную и связанную.
Свободная-играет рольрастворителя и как дисперсионная коллоидная система.
Связанная- образует слабые связи- водородная, вандервальсовые с полярными группами разных молекул.
Связанная и свободная вода различается по подвижности молекул воды и растворяющей способности. Наименьшей подвижностью и растворяющей способностью обладает внутренняя связанная вода. Она присутствует в липидной зоне мембран в виде отдельных молекул. Основную часть связанной воды представляет вода гидратных оболочек. Эта вода окружает полярные группы белков и липидов, имеет min подвижность и практически не обладает свойствами растворителя. Свободная вода в порах и каналах. По ней могут перемещаться свободные ионы. Она является хорошим растворителем, подвижная и обладает всеми свойствами жидкой воды.
Захваченная вода обладает изотопным движением, характерным для жидкой воды, является хорошим растворителем. Она встречается в центральной зоне мембран, между ее липидными слоями, но эта вода пространственно делится как с внеклеточной жидкостью, так и с цитоплазмой. У нее нет возможности свободно с ними обмениваться.
Структурные гипотезы: 1) Квадикристаллическая – данная модель предполагает, что длина связей и расстояние соседних молекул одинаково.
2) Микрокристаллическая - в жидкости существуют области, разделенные четкими границами. Эти области называют кластерами. Они разделены пуст-ми из соверш-но свобод-х молекул. М/у кластерами и пустотами происходит постоянный обмен. 70 % воды наход-ся в виде класстеров, 30 % в виде пустот. С увелич темпер кол-во класстеров и их размеры уменьш.
11. Адгезия живых клеток
Одна из важнейших функций плазматических мембран — обеспечение сцепления клеток друг с другом (адгезия). Соединение клеток возможно благодаря наличию на их поверхности специализированных структур — выростов. Осуществляется сцепление клеток разными способами: в одних случаях создается механическое сцепление по типу «выступ — гнездо», в других между молекулами, входящими в состав структур-выступов, устанавливается химическая связь. Существует предположение, что межклеточная жидкость содержит склеивающее вещество. Основой для такого типа контакта может быть органическая соль кальция, образующая связи с карбоксильными группами белков и липидов. Это предположение подтверждается тем, что с понижением концентрации кальция в межклеточной жидкости способность клеток к адгезии уменьшается.
Взаимодействие клеток существенно зависит от величины поверхностного заряда ПМ. Отрицательный заряд на поверхности клеток вызывает их взаимное отталкивание, сила которого определяется величиной электрокинетического потенциала. Если действием трипсина вызвать полную дезинтеграцию клеток тканей (например, куриного эмбриона), то через некоторое время можно наблюдать слияние однотипных клеток и образование агрегатов из клеток одной ткани. Если смешать клетки зародышей разных видов, то их последующая адгезия будет связана не с видовой специфичностью, а с тканевой принадлежностью. По-видимому, организация клеток в ткани обусловлена локализацией на клеточной поверхности специального механизма. Способность к адгезии у всех клеток различна: клетки злокачественных опухолей обладают более низкой способностью к адгезии, чем нормальные, что объясняется пониженным содержанием Са2+ в раковых клетках и соответственно их высоким поверхностным зарядом. Повышение электрокинетического потенциала вызывает возрастание сил взаимного отталкивания, поэтому опухолевые клетки обладают более высокой подвижностью по сравнению с нормальными. Они легко отрываются от опухоли и уносятся с током тканевой жидкости, инициируя образование метастазов.
Клеточную адгезию реализуют специальные гликопротеины — молекулы адгезии. Прикрепление клеток к компонентам внеклеточного матрикса осуществляют точечные (фокальные) адгезионные контакты, а прикрепление клеток друг к другу — межклеточные контакты. Молекулы адгезии специфичны для каждого типа ткани. Так, Е-кадгерин связывает клетки эмбриональных тканей, Р-кадгерин — клетки плаценты и эпидермиса, N-CAM — клетки нервной системы и т.д. Адгезия позволяет клеточным партнёрам обмениваться информацией ч/з сигнальные молекулы плазматических мембран и щелевые контакты.
Различают 2 гр мол-л адгезии: Кадгерины — трансмембранные гликопротеины нескольких типов. Надсемейство иммуноглобулинов вклю-т несколько форм молекул адгезии нервных клеток — (N-CAM), молекулы адгезии L1, нейрофасцин и др. Экспрессируются в нервной ткани.
Адгезионный контакт. Прикрепление клеток к молекулам адгезии внеклеточного матрикса реализуют точечные (фокальные) адгезионные контакты. Адгезионный контакт содержит винкулин, α-актинин, талин и другие белки. В образовании контакта участвуют также трансмембранные рецепторы — интегрины, объединяющие внеклеточные и внутриклеточные структуры. Характер распределения макромолекул адгезии во внеклеточном матриксе (фибронектин, витронектин) определяет место окончательной локализации клетки в формирующейся ткани.