
Последствия для эволюции и не только








АРХИТЕКТУРА ЖИЗНИ
Дональд Э. Ингбер
«По-видимому, построение органических структур — от простых углеродных соединений до сложных клеток и тканей — определяет универсальный набор правил».
Что такое Тенсегрити?
От скелета к цитоскелету
Жёсткая проводка в клетках (ячейках)
Как механика управляет биохимией
Заставить клетки сделать поворот
Молекулярные геодезические купола
Универсальный шаблон
Последствия для эволюции и не только
ИЛЛЮСТРАЦИИ:
Архитектура цитоскелета
Линейное усиление
Трансформация Тенсегрити (Quicktime)
Модель Тенсегрити (VRML)
ДАЛЬНЕЙШЕЕ ЧТЕНИЕ
ССЫЛКИ ПО ТЕМЕ
Большинство исследователей через некоторое время признают тот интригующий факт, что большинство из них игнорируют в своих устремлениях объяснение основ жизни.
Жизнь имеет высшую сложность функционирования. Организм, будь то бактерия или бабуин, развивается через невероятно сложную серию взаимодействий, включающую огромное количество различных компонентов. Эти компоненты или подсистемы сами состоят из более мелких молекулярных компонентов, которые независимо демонстрируют свое динамическое поведение, например, способность катализировать химические реакции. Однако, когда они объединяются в некую более крупную функциональную единицу, такую, как клетка или ткань, появляются совершенно новые и непредсказуемые свойства, в том числе способность двигаться, изменять форму и расти.
Биологи в течение последних нескольких десятилетий пытались углубить наше понимание того, как работает человеческое тело, определяя свойства жизненно важных материалов и молекул, таких, как ДНК - материал для генов. Сейчас биологи стремятся идентифицировать каждый ген в полном наборе, известном как геном, который несёт каждый человек. Ведь, поскольку гены являются «чертежами» для ключевых белков, этот «Святой Грааль» молекулярной биологии в ближайшем будущем приведёт к созданию каталога практически всех молекул, из которых создан человек. Однако понимание того, из чего состоят части сложной машины, мало что даёт для объяснения того, как работает вся система, независимо от того, является ли сложная система двигателем внутреннего сгорания или ячейкой (эмерджентость, или системные свойства, см. выше). Другими словами, определение и описание кусочков молекулярной головоломки мало что даст, если мы не понимаем правил их сборки.
Несмотря на описанную сложность, природа применяет некоторые общие правила сборки, что видно из применения и повторения определённых шаблонов всего в масштабах от молекулярного до макроскопического, таких, как спирали, пятиугольники и треугольные формы. Эти шаблоны проявляются в структурах, варьирующихся от очень регулярных кристаллов до относительно нерегулярных белков и даже в таких разнообразных организмах, как вирусы, планктон и люди. Хотя и органические, и неорганические вещества состоят из одних и тех же строительных блоков в виде атомов углерода, водорода, кислорода, азота и фосфора, и единственная разница заключается в том, как эти атомы расположены в трёхмерном пространстве.
Явление, при котором компоненты соединяются вместе, образуя более крупные устойчивые структуры, обладающие новыми свойствами, которые нельзя было предсказать на основании характеристик их отдельных частей, известно как самосборка. Она наблюдается во многих масштабах в природе. В человеческом теле, например, большие молекулы самособираются в клеточные компоненты, известные как органеллы, которые в свою очередь самособираются в клетки, а те самособираются в ткани, последние самособираются в органы и т.д., результатом чего является тело, организованное иерархически как уровни систем внутри систем, и т.д. в надорганизменных уровнях. Таким образом, если мы хотим полностью понять, как формируются и функционируют живые существа, нам необходимо раскрыть эти основные принципы, которыми руководствуется биологическая организация.
Несмотря на столетия исследований, исследователи до сих пор относительно мало знают о силах, которые направляют атомы на самосборку в молекулы. Еще меньше они знают о том, как группы молекул объединяются для создания живых клеток и тканей. Однако за последние два десятилетия я обнаружил и исследовал интригующий и, казалось бы, фундаментальный аспект самосборки: поразительно большое разнообразие природных систем, включая атомы углерода, молекулы воды, белки, вирусы, клетки, ткани и даже люди и другие живые существа построены с использованием распространенной ныне формы архитектуры, известной как тенсегрити. Этот термин относится к системе, которая механически стабилизируется благодаря тому, как силы растяжения и сжатия распределяются и уравновешиваются внутри конструкции.
Это фундаментальное открытие может однажды найти практическое применение во многих областях. Например, новое понимание тенсегрити на клеточном уровне позволило нам лучше понять, как форма клеток и механические силы, такие как давление в кровеносных сосудах или сжатие в костях, влияют на активность генов. В то же время более глубокое понимание естественных правил самосборки позволит нам лучше использовать — в различных приложениях от разработки лекарств до тканевой инженерии — быстро накапливающиеся данные о молекулах, клетках и других биологических компонентах. Объяснение того, почему тенсегрити так распространено в природе, может также дать новый взгляд на те самые силы, которые управляют биологической организацией, и, возможно, на саму эволюцию.
Что такое Тенсегрити?
Мой интерес к тенсегрити восходит к моим студенческим годам в середине 1970-х годов в Йельском университете. Там мои исследования клеточной биологии, а также скульптуры привели меня к пониманию того, что вопрос о том, как формируются живые существа, связан не столько с их химическим составом, сколько с архитектурой. Исходя из того факта, что молекулы и клетки наших тканей постоянно удаляются и заменяются; я рассудил, что поддержание шаблона и архитектуры мы и называем жизнью.
Тенсегрити-структуры механически устойчивы не из-за прочности отдельных элементов, а из-за того, как вся конструкция распределяет и уравновешивает механические напряжения. Такие структуры делятся на две категории.
Конструкции одной категории, в которую входят геодезические купола Бакминстера Фуллера, в основном представляют собой каркасы, состоящие из жёстких стоек, каждая из которых может выдерживать растяжение или сжатие. Стойки, составляющие каркас, соединены в треугольники, пятиугольники или шестиугольники, то есть каждая стойка ориентирована таким образом, чтобы зафиксировать каждое соединение в фиксированном положении, тем самым обеспечивая устойчивость всей конструкции.
Другая категория структур тенсегрити включает в себя те, которые стабилизируются посредством явления, известного как предварительное напряжени. ещё до того, как одна из этих конструкций подвергнется воздействию внешней силы, все элементы конструкции уже находятся в состоянии растяжения или сжатия, т. е. предварительно напряжены. Этот тип конструкции был впервые построен скульптором Кеннетом Снельсоном. В элегантных скульптурах Снельсона структурные элементы, которые могут выдерживать только растяжение, отличаются от тех, которые выдерживают сжатие в конструкциях первого типа, при этом внутри конструкции отдельные жёсткие стойки, несущие сжатие, растягивают (натягивают) гибкие растянутые элементы (тросы), и в то же время эти несущие элементы сжимают жёсткие стойки – таким образом, эти противодействующие силы, уравновешивающие всю структуру, позволяют ей стабилизироваться.
Структуры тенсегрити обеих категорий имеют одну важную особенность: напряжение постоянно передается сразу на все структурные элементы, другими словами, увеличение напряжения в одном из элементов приводит к увеличению напряжения сразу во всех элементах конструкции (даже на противоположной стороне). В структурах второго типа глобальное увеличение натяжения (растяжения) уравновешивается увеличением сжатия элементов, разнесённых (распределённых) по всей структуре, и наоборот - таким образом, эти структуры стабилизируются с помощью механизма, который Фуллер описал как непрерывное растяжение и локальное сжатие. Напротив, устойчивость большинства зданий обеспечивается постоянным сжатием под действием внешней (не внутренней) силы тяжести.
Несущие элементы в обеих типах структур, будь то купола Фуллера или скульптуры Снельсона, намечают кратчайшие пути между соседними элементами и, следовательно, по определению расположены геодезически, поскольку силы натяжения естественным образом передаются по кратчайшему расстоянию между двумя точками. Поэтому элементы тенсегрити-структур располагаются точно так, чтобы наилучшим образом противостоять нагрузкам, а сами эти структуры обеспечивают максимально возможную прочность для данного количества строительного материала.
От скелета к цитоскелету
Какое отношение принцип тенсегрити имеет к человеческому телу? Применение принципа тенсегрити наюлюдаются практически во всех обнаруживаемых масштабах тела. Так, на макроскопическом уровне все 206 костей, составляющих наш скелет, вытягиваются против силы гравитации и стабилизируются в вертикальной форме натяжением растянутых мышц, сухожилий и связок (аналогично тросам в скульптурах Снельсона). Другими словами, в сложной тенсегрити-структуре внутри каждого из нас кости являются распорками сжатия, а мышцы, сухожилия и связки — несущими элементами. С другой стороны, как оказалось, белки и другие ключевые молекулы в организме также стабилизируются с помощью принципов тенсегрити. Область моих собственных интересов лежит между этими двумя крайностями - на клеточном уровне.
Будучи аспирантом, работавшим с Джеймсом Д. Джеймисоном в Йельском университете, я сосредоточился на понимании того, как компоненты биологических систем (особенно клеток) взаимодействуют механически. В то время, в конце 1970-х биологи обычно рассматривали клетку просто как вязкую жидкость или гель, окружённый мембраной, что очень похоже на воздушный шар, наполненный патокой. Хотя уже было известно, что клетки содержат внутренний каркас - цитоскелет, состоящий из трёх различных типов молекулярных белковых полимеров, известных как микрофиламенты, промежуточные филаменты и микротрубочки, их роль в контроле формы клеток была плохо изучена.
Ещё одна загадка тех дней касалась поведения изолированных клеток, помещённых на разных поверхностях. Было давно известно, что клетки расширяются и уплощаются, когда они прикрепляются к чашке из жёсткого стекла или пластика. Однако в 1980 году Альберт К. Харрис из Университета Северной Каролины в Чапел-Хилл показал, что при прикреплении к гибкому каучуковому субстрату клетки, наоборот, сжимаются и становятся более сферическими, а это сокращение в свою очередь сжимает (сморщивает) нижележащую поверхность. И мне пришло в голову, что взгляд на клетку как на тенсегрити-структуру может легко объяснить такое поведение. Я смоделировал клетку как такую структуру; состоящую из шести деревянных стоек и эластичной нити, расположив стойки, выдерживающие сжимающее напряжение, тремя парами: каждая пара была перпендикулярна двум другим, и ни одна из деревянных распорок фактически не касалась друг друга. При этом эластичная нить, находящаяся под натяжением, была соединена с концами всех стоек, придавая им устойчивую трёхмерную форму. Я также поместил меньшую сферическую тенсегрити-модель, моделирующую ядро, внутрь более крупной, которая представляла остальную часть клетки. Затем, чтобы имитировать связи цитоскелета между ядром и остальной клеткой, я протянул эластичные нити от поверхности большой тенсегрити-структуры к меньшей внутри.
Чтобы понять, как работал мой эксперимент, необходимо знать, что при нажатии на тенсегрити-модель того типа, который я построил, она превращается в сплющенную груду палочек и веревок. Но как только давление снимается, энергия, накопленная в натянутых нитях, заставляет модель возвращаться к своей первоначальной, примерно сферической форме. Чтобы смоделировать поведение клеток при размещении на поверхности, я имитировал твёрдый культуральный субстрат из стекла или пластика, натянув кусок ткани и крепко прикрепив его булавками к дереву под ним, и прикрепил тенсегрити-модель к этой основе, предварительно сплющив её и пришив концы некоторых стоек к ткани - эти прикрепления были аналогичны молекулам клеточной поверхности (мембраны), теперь известным как интегрины, или рецепторы адгезии, которые физически соединяют клетку с её якорным субстратом.
При пришивании концов стоек к плотно сколоченной ткани модель оставалась плоской, как настоящая клетка на твёрдой подложке, однако, когда я поднял булавки, чтобы освободить ткань от дерева, тем самым сделав поверхность крепления ячейки гибкой, тенсегрити-модель сразу выскочила в более сферическую форму, сморщив ткань под собой. Кроме того, я заметил, что, когда я растягивал модель, соединяя её с тканевой подложкой, клетка и ядро внутри неё расширялись скоординированным образом (совместно), при этом модель ядра также сместилась к нижней части моделируемой ячейки. Вскоре после этого я экспериментально показал, что живые клетки и ядра деформируются и поляризуются сходным образом, когда прилипают к субстрату - таким образом, с помощью моей очень упрощённой конструкции я показал, что тенсегрити-структуры имитируют известное поведение живых клеток.
Жёсткая проводка в клетках
За годы, прошедшие после моего модельного эксперимента, биологи многое узнали о механических аспектах клеток, и их открытия, кажется, подтверждают, что клетки действительно получают и сохраняют свою форму благодаря тенсегрити. Далее, как и предсказывают модели, большинство клеток получают свою структуру не только из трёх основных типов филаментов цитоскелета, но и из внеклеточного матрикса — якорного каркаса, к которому клетки естественным образом крепятся в организме.
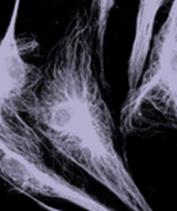
Архитектура цитоскелета
Внутри клетки тонкая сеть сократительных микрофиламентов — ключевой элемент цитоскелета — простирается по всей клетке, оказывая натяжение, то есть притягивая клеточную мембрану и все её внутренние компоненты к ядру в сердцевине. Этому внутреннему натяжению противостоят два основных типа сжимающих элементов, один из которых находится вне клетки, а другой – внутри: компонент вне клетки представляет собой внеклеточный матрикс, а сжатые «балки» внутри клетки могут быть либо микротрубочками, либо большими пучками сшитых микрофиламентов внутри цитоскелета. Третий компонент цитоскелета - промежуточные филаменты - являются крупными интеграторами, соединяющими микротрубочки и сократительные микрофиламенты друг с другом, а также с поверхностной мембраной и ядром клетки. Кроме того, они действуют как растяжки, укрепляя центральное ядро и фиксируя его на месте. И хотя цитоскелет окружен гибкими мембранами и пронизан вязкой жидкостью, эта жёсткая сеть молекулярных распорок и тросов вполне стабилизирует форму клетки.
Но если клетка и ядро физически связаны натяжными нитями, а не только жидкой цитоплазмой, то натяжение рецепторов на поверхности клетки должно вызывать немедленные структурные изменения глубоко внутри клетки. Недавно Эндрю Маниотис, который был в моей группе в Детской больнице Гарвардской медицинской школы, прямо продемонстрировал это: связывая микропипетки с рецепторами адгезии на поверхности живых клеток и вытягивая их наружу, он вызывал немедленную перестройку цитоскелетных филаментов и структур в ядре в направлении натяжения. Таким образом, как показали и мои ранние эксперименты, клетки и ядра не ведут себя подобно вязким воздушным шарам с водой.
Как механика управляет биохимией
Однако, принцип тенсегрити может объяснить большее, чем просто стабилизацию формы клеток и ядер. Стивен Р. Хайдеманн, работая с Харишем Джоши и Робертом Э. Буксбаумом из Мичиганского государственного университета, в середине 1980-х годов обнаружил, что тенсегрити может объяснить, как нервные клетки вытягивают длинные тонкие выступы, называемые нейритами, которые заполнены микротрубочками и передают электрические сигналы в нервной системе - этот рост необходим для восстановления повреждённого нерва. Группа Хайдемана обнаружила, что микротрубочки сжимаются на своих концах за счёт притяжения окружающих сократительных микрофиламентов внутри нейритов. Что ещё более важно, исследователи обнаружили, что сборка микротрубочек (удлинение) и, следовательно, расширение нейритов обеспечивается за счёт смещения сжимающих нагрузок с микротрубочек на места прикрепления клетки к её внеклеточному матриксу. Совсем недавно Эндрю Матус из Института Фридриха Мишера в Базеле добавил к этой истории яркое примечание: создавая клетки, производящие флуоресцентные микротрубочки, Матус фактически наблюдал, как эти микротрубочки изгибаются при сжатии.
Другими словами, существование баланса сил тенсегрити даёт возможность интегрировать на молекулярном уровне механику и биохимию. Модель тенсегрити предполагает, что структура цитоскелета клетки может быть изменена путём изменения баланса физических сил, передаваемых через клеточную поверхность. Это открытие очень важно, поскольку многие ферменты и другие вещества, контролирующие синтез белка, преобразование энергии и рост клетки, физически иммобилизованы на цитоскелете - по этой причине изменение геометрии и механики цитоскелета может влиять на биохимические реакции и даже изменить активируемые гены и, следовательно, синтезируемые белки.
Для дальнейшего изучения этой возможности Рахул Сингхви и Кристофер С. Чен из моей группы, работая с Джорджем М. Уайтсайдсом из Гарварда, разработали метод изменения формы и функции клеток: они заставляли живые клетки принимать различные формы — сферические или уплощенные, круглые или квадратные — просто помещая их на крошечные липкие «островки», состоящие из внеклеточного матрикса, при этом каждый адгезивный островок был окружен тефлоновой поверхностью, к которой клетки не могли прилипать. И этими простыми изменениями формы клетки они могли переключать из различные генетические программы: например, плоские клетки делились с большей вероятностью, тогда как круглые клетки, которым не давали двигаться, активировали программу смерти, известную как апоптоз; когда же клетки не были ни слишком вытянуты, ни слишком втянуты, они не делились и не умирали, а вместо этого дифференцировались тканеспецифическим образом: капиллярные клетки образовывали полые капиллярные трубки, клетки печени секретировали белки, которые печень обычно поставляет в кровь, и так далее.
Таким образом, механическая перестройка клетки и цитоскелета, по-видимому, говорит клетке, что делать: очень плоские клетки с растянутым цитоскелетом чувствуют, что нужно больше клеток, чтобы покрыть окружающий субстрат — как при заживлении ран — и, следовательно, необходимо деление клеток. Округление же клеток указывает на то, что за место в матрице борются слишком много клеток и они слишком быстро размножаются; поэтому некоторые должны умереть, чтобы предотвратить образование опухоли. Нормальная функция тканей устанавливается и поддерживается между этими двумя крайностями.
Понимание того, как происходит это переключение, может привести к новым подходам в терапии рака и восстановлению тканей и, возможно, даже к созданию искусственных заменителей тканей.
Заставить клетки сделать поворот
Следующим уровнем в иерархии самосборки является образование тканей, которые создаются путём соединения клеток друг с другом и с их внеклеточным матриксом. Одним из возникающих при этом свойств тканей является то, как они ведут себя механически: многие различные типы тканей - включая мышцы, хрящи, кровеносные сосуды и кожу - проявляют реакцию, известную как линейное уплотнение. Например, если вы потянете кожу, вы почувствуете, что сопротивление увеличивается по мере того, как вы тянете сильнее - возрастающая внешняя сила встречает возрастающее сопротивление. Недавние исследования показали, что линейную жёсткость проявляют даже изолированные молекулы, такие как ДНК. Однако механического или математического объяснения такого типа поведения не было, пока мы не исследовали это явление в контексте тенсегрити.
В 1993 году мой коллега Нин Ван, работая с Джеймсом П. Батлером из Гарвардской школы общественного здравоохранения, разработал устройство, которое позволяло нам скручивать отдельные молекулы на поверхности мембраны живых клеток, одновременно измеряя клеточный ответ. Мы обнаружили, что когда мы увеличивали нагрузку на интегрины (молекулы, которые проходят через клеточную мембрану и связывают внеклеточный матрикс с внутренним цитоскелетом), клетки реагировали, становясь всё более и более жёсткими, как целые ткани. Кроме того, живые клетки можно делать жёсткими или гибкими, изменяя предварительное напряжение в цитоскелете - например, при изменении натяжения сократительных микрофиламентов.

Линейное усиление
Хотя не все детали взаимодействия точно известны, мы показали, используя модель тенсегрити со стойками и нитью, что суть этой реакции можно объяснить подобно тому, как структуры тенсегрити реагируют на стресс (локальную нагрузку): по сути, в ответ на локальное напряжение все взаимосвязанные структурные элементы модели тенсегрити перестраиваются. При этом линейное усиление возникает из-за того, что по мере увеличения приложенного напряжения всё больше стержней (стоек) выстраиваются в направлении приложенного напряжения.
Работая с Дмитрием Стаменовичем из Бостонского университета, мы разработали математическую модель, основанную на этих принципах. Она впервые предсказывает линейную реакцию жёсткости тканей, живых клеток и даже молекул. Мы надеемся использовать эту модель при разработке передовых материалов, обладающих свойством линейной жесткости, которые могут быть полезны в таких приложениях, как защитная одежда и изготовление протезов. Тот же математический подход можно использовать в компьютерных программах для ускорения моделирования молекулярной динамики при разработке лекарств.
При исследованиях магнитного скручивания Ванга и в экспериментах Маниотиса с вытягиванием микропипеток мы обнаружили, что приложение стресса к рецепторам клеточной поверхности, участвующим в метаболизме, а не в адгезии, не обеспечивает эффективной передачи силы внутрь клетки - таким образом, эти исследования подтвердили, что механические силы передаются лишь по определённым молекулярным путям в живых клетках, что дало новое понимание того, как клетки воспринимают механические стимулы, регулирующие развитие тканей. Это понимание, в свою очередь, может помочь нам лучше понять широкий спектр явлений - от роста мышц в ответ на напряжение до роста корней растений в присутствии гравитации.
Молекулярные геодезические купола
Хотя модели тенсегрити предсказывали поведение многих клеток, требовало объяснения одно проявившееся несоответствие: многие клетки могут расширяться и сплющиваться без микротрубочек — наиболее важных сжимающих распорок в модели, и если живые клетки могут превращаться из сферических в плоские без этих распорок, то зачем применять тенсегрити? Однако, опять же, используя несложный подход к моделированию, я обнаружил, что сеть микрофиламентов сама по себе представляет собой тенсегрити-структуру: в цитоскелете живой клетки сократительные микрофиламенты образуют решётку, которая локально реорганизуется в различные формы, такие, как большие пучки или сети треугольников.
Чтобы изучить механизм этой реорганизации, я смоделировал решётку микрофиламентов в виде многогранного каркаса из соломинок для газированных напитков, который содержал шесть треугольников и четыре квадрата. Соломинки были скреплены одной эластичной нитью, которую я пропустил через все соломинки и привязал к себе. При этом я предполагал, что каждая соломинка в модели представляет собой единственную сократительную микронить, которая может генерировать механическое напряжение, укорачивая себя. Известно, что сократительные микрофиламенты при укорочении становятся жёстче - таким образом, внутренняя эластичная нить в модели будет имитировать постоянное напряжение во всей структуре, возникающее в результате укорочения всех этих затвердевших нитей.

Трансформация тенсегрити
При этом я предположил, что такая модель представляет собой одну модульную единицу цитоскелета, которая во всех направлениях связана с другими подобными модулями в круглой (неприкрепленной, а подвешенной) клетке. И я пытался ответить на вопрос, что произойдет с этим каркасом, если поддерживающая его клетка прикрепится к жёсткой поверхности?
Клетки прикрепляются, связываясь с поверхностно-связанными молекулами внеклеточного матрикса, причём эти клетки «приклеены» к матрице неравномерно, даже, скорее, они «точечно сварены» в локализованных местах, известных как фокальные спайки. Сократительные микрофиламенты реагируют на это закрепление укорачиванием и увеличением изометрического напряжения внутри решётки. В модели из содовой соломки предполагалось, что увеличивающееся напряжение, вызванное прикреплением, заставит отдельные сократительные микрофиламенты, которые в модели сформировали квадраты, самособираться в линейные пучки, протянувшиеся между этими участками фокальной адгезии, где интегриновые рецепторы прикрепляют клетку к матрице. И, на самом деле, оказалось, что когда живые клетки распространяются по поверхности, то отдельные сократительные микрофиламенты выстраиваются почти идентичным образом, образуя пучки, называемые стрессовыми волокнами.
Напротив, поскольку в верхней части клетки нет адгезивной подложки, которая противостояла бы натяжению укорачивающих микрофиламентов, то в этих областях сокращению каждого микрофиламента может сопротивляться только натяжение и жёсткость соседних филаментов. Фуллер много лет назад показал, что внутреннее растяжение и скручивание заставляют этот тип многогранной структуры подвергаться так называемой джиттербаг-трансформации: очень гибкий каркас из квадратов и треугольников превращается в полностью треугольные октаэдрические или тетраэдрические формы, или, другими словами, в полностью триангулированные структуры тенсегрити. Когда я соединил между собой множество похожих моделей из соломки, то обнаружил, что отдельные модули постепенно сжимаются, что приводит к образованию геодезического каркаса, состоящего из чередующихся октаэдрических и тетраэдрических форм, плотно упакованных. И сокращение в клетке окружающих сетей микрофиламентов, которые взаимодействуют с клеточным основанием, изгибало бы этот каркас вниз над сферическим ядром, тем самым превращая его в почти трёхгранный купол (в частности, в геодезический купол). Элиас Лазаридес, тогда работавший в лаборатории Колд-Спринг-Харбор в Нью-Йорке, а также Мэри Осборн и Клаус Вебер из Института Макса Планка в Геттингене, Германия, наблюдали именно эти преобразования в области цитоплазмы над ядром в распространяющихся клетках. Примечательно, что существование геодезического купола внутри цитоскелета на молекулярном уровне убедительно демонстрирует, что клетки могут использовать и действительно используют архитектуру тенсегрити для формирования своего цитоскелета.
Универсальный шаблон
Геодезическая структура, обнаруженная в цитоскелете, является классическим примером шаблона, который встречается в природе повсюду во многих масштабах. Сферические группы атомов углерода, называемые бакминстерфуллеренами, или бакиболлами, наряду с вирусами, ферментами, органеллами, клетками и даже небольшими организмами имеют геодезические формы. И, как ни странно, лишь немногие исследователи, похоже, задавались вопросом, почему это так. Я считаю, что этот повторяющийся шаблон является визуальным свидетельством существования общих правил самосборки. В частности, все эти сущности стабилизируют себя в трёх измерениях схожим образом - упорядочивая свои части так, чтобы минимизировать энергию и массу, за счёт общего натяжения и локального сжатия, то есть за счёт тенсегрити.
Например, сборка вирусов - мельчайшей формы жизни на Земле - включает связывающие взаимодействия между многими схожими белками, которые объединяются, чтобы сформировать геодезическую вирусную оболочку, которая заключает в себе генетический материал. Во время формирования вируса линейные отростки белков перекрываются с аналогичными хвостами, которые отходят от соседних белков, образуя триангулированный геодезический каркас в нанометровом масштабе. Каждое соединение в этой структуре самостабилизируется в результате баланса межмолекулярных сил притяжения (водородные связи) и способности отдельных белковых хвостов сопротивляться сжатию и изгибу.
Та же основная схема проявляется и в бакиболах, лишь с тем отличием, что строительными блоками здесь являются атомы, а не белки: в бакиболах 60 атомов углерода образуют геодезическую сферу, покрытую 20 шестиугольниками, перемежающихся 12 пятиугольниками – подобно узору на футбольном мяче. При этом 90 углерод-углеродных связей в бакиболе, по сути, являются распорками тенсегрити.
Труднее увидеть, что те же правила построения применимы и к неправильным (асимметричным) структурам, включая многие биологические молекулы, которые не имеют геодезических форм. Белки, от которых зависит структура клеток, катализ и многие другие функции, представляют собой длинные цепочки аминокислот. При этом небольшие участки аминокислотного остова белка складываются в спиралевидные формы, которые стабилизируются за счёт баланса между силой притяжения водородных связей (стягивающих вместе различные области молекулы) и способностью белкового витка сопротивляться укорочению или сжатию - другими словами, эти спиральные области стабилизируются также за счёт тенсегрити, как и любая спиральная молекула, например, ДНК.
Белковая организация включает также иерархическую сборку: небольшие области белка, имеющие спиральную жёсткость, отделены друг от друга частями одного и того же аминокислотного остова, которые действуют подобно гибким шарнирам. Эти стержнеобразные области складываются сами по себе (из-за сил растяжения водородных связей), чтобы стабилизировать всю молекулу. Жёсткие спирали могут быть сильно сжаты локально, даже если силы уравновешены по всей предварительно напряжённой молекуле, а поскольку локальная сила может изменить форму всей структуры тенсегрити, то связывание молекулы с белком может привести к тому, что различные затвердевшие спиральные области изменят своё относительное положение по всей длине белка: например, когда молекула, несущая сигнал, связывается с рецептором, который проходит через мембрану и проникает в клетку, то это присоединение может вызывать конформационные изменения на противоположном конце рецептора, что в свою очередь изменяет форму соседних белков и запускает каскад молекулярной реструктуризации внутри клетки. И действительно, клетки чувствуют и реагируют на изменения в окружающей их среде подобным образом.
Таким образом, тенсегрити - от молекул до костей, мышц и сухожилий человеческого тела - явно является предпочтительной строительной системой природы. Только тенсегрити, например, может объяснить, почему каждый раз, когда вы двигаете рукой, ваша кожа натягивается, а при расширении внеклеточного матрикса ваши клетки искажаются, и взаимосвязанные молекулы во внутренней структуре клетки ощущают притяжение — и всё это без действий вроде их поломок, или разрывов.
Примечательно, что принцип тенсегрити даже может объяснить, почему все эти явления так идеально согласованы в живом существе. В Медицинской школе Джона Хопкинса Дональд С. Коффи и Кеннет Дж. Пьента обнаружили, что структуры тенсегрити функционируют как связанные гармонические осцилляторы, поскольку ДНК, ядра, нити цитоскелета, мембранные ионные каналы и целые живые клетки и ткани обладают характерными резонансными частотами колебаний. Проще говоря, передача напряжения через массив тенсегрити обеспечивает как средство распределения усилий по всем взаимосвязанным элементам, так и соединение с «настройкой» всей системы как единого механического целого.
Последствия для эволюции и не только
Хотя биологическое разнообразие порождают изменения в ДНК, гены являются лишь продуктом эволюции, а не её движущей силой. На самом деле, геодезические формы, подобные тем, которые обнаружены у вирусов, ферментов и клеток, существовали в неорганическом мире кристаллов и минералов задолго до появления ДНК. Даже молекулы воды имеют геодезическую структуру. Тогда уместен вопрос, как органические молекулы и клетки произошли от неорганических компонентов? Ведь, в конце концов, с точки зрения возниковения эмерджентных (системных, интегральных) свойств, самосборка молекул в органеллы или клеток в ткани не сильно отличается от самосборки атомов в соединения. Например, натрий - взрывоопасный металл, и хлор - ядовитый газ - объединяются с образованием хлорида натрия, чьё новое свойство заключается в том, что его уже можно использовать в качестве поваренной соли. Таким образом, важным принципом здесь является то, как структура формирует себя и удерживает вместе свои компоненты в трёхмерном пространстве – именно эта характеристика определяет, как будет вести себя структура в целом.
В более широком смысле, одним и тем же пространственным ограничениям подчиняется вся материя, независимо от её масштаба или положения. С учётом этого, поскольку принцип тенсегрити является наиболее экономичным и эффективным, он может использоваться как способ построения в молекулярном, макроскопическом и во всех промежуточных масштабах. Вполне возможно, что полностью триангулированные тенсегрити-структуры были выбраны в ходе эволюции из-за их структурной эффективности — высокой механической прочности при использовании минимума материалов, вместе с гибкостью, демонстрируемой предварительно напряжёнными структурами тенсегрити, что выгодно, поскольку позволяет конструкциям принимать самые различные формы. Например, если молекула или клетка могут трансформироваться в более устойчивую форму лишь при определённой температуре или давлении или в более эффективную в метаболическом отношении форму (как это и имеет место в действительности), то срок их жизни может быть продлён и более вероятно их взаимодействие с другими подобными сущностями, а затем они снова могут собираться самостоятельно.
Теперь исследователи считают, что биологическая эволюция началась в слоях глины, а не в первозданном море (с учётом тоже лечебных её свойств, как и у морской воды), поскольку глина представляет собой пористую сеть атомов, как раз расположенных геодезически в октаэдрических и тетраэдрических формах. Но поскольку эти октаэдры и тетраэдры не плотно упакованы, то они сохраняют способность двигаться и скользить относительно друг друга - эта гибкость, по-видимому, и позволяет глине катализировать многие химические реакции, в том числе те, которые могли привести к образованию первых молекулярных строительных блоков органической жизни. Со временем различные молекулярные коллективы самособирались, образуя первые структуры со специализированными функциями — предшественники современных органелл, которые затем объединялись друг с другом, создавая первые простые клетки. Затем эти клетки продуцировали белки, которые самособирались с образованием внеклеточного матрикса — «заякоренных» каркасов, которые в свою очередь способствовали самосборке многоклеточных тканей. Органы же развились из самосборки тканей, а сложные организмы - возникли в результате комбинирования и постепенного ремоделирования различных органов. И действительно, развитие эмбриона из сперматозоида и яйцеклетки повторяет все эти этапы самосборки.
Хотя появление ДНК и генов породило новый механизм создания структурного разнообразия, ускоривший эволюцию, однако на протяжении всего этого времени правила, управляющие процессом иерархической самосборки, оставались по существу неизменными. Поэтому неудивительно, что базовое расположение костей и мышц у Tyrannosaurus rex и Homo sapiens удивительно похоже; что животные, насекомые и растения полагаются на предварительное напряжение для механической устойчивости своих тел; и что в природных системах преобладают геодезические формы, такие, как шестиугольники, пятиугольники и спирали.
Наконец, возникают более философские вопросы, универсальны ли эти строительные принципы и применимы ли они к структурам, формируемым как очень крупными, так и малыми силами? Мы не знаем. Снельсон, однако, предложил интригующую модель атома, основанную на тенсегрити, которая начинается там, где французский физик Луи де Бройль остановился в 1923 году. Сам Фуллер при этом зашёл так далеко, что представил Солнечную систему как структуру, состоящую из множества недеформируемых колец планетарного движения, поддерживаемую постоянным гравитационным напряжением. Однако, тот факт, что наша расширяющаяся (напрягающаяся) Вселенная содержит огромные нити гравитационно связанных галактик и изолированных чёрных дыр, которые локально испытывают огромные сжимающие силы, не может не вызывать у нас удивления – поэтому, вполне возможно, что у природы действительно задана лишь одна основная тема. Как предположил шотландский зоолог начала 20-го века Д'Арси В. Томпсон, который цитировал Галилея, а тот в свою очередь цитировал Платона: Книга Природы действительно может быть написана геометрическими буквами.
ДАЛЬНЕЙШЕЕ ЧТЕНИЕ
ON GROWTH AND FORM. REVISED EDITION. D'Arcy W. Thompson. Cambridge University Press, 1942 (reprinted 1992).
MOVEMENT AND SELF-CONTROL IN PROTEIN ASSEMBLIES. Donald L. D. Caspar in Biophysical Journal, Vol. 32, No. 1, pages 103--138; October 1980.
CLAY MINERALS AND THE ORIGIN OF LIFE. Edited by A. Graham Cairns-Smith and Hyman Hartman. Cambridge University Press, 1986.