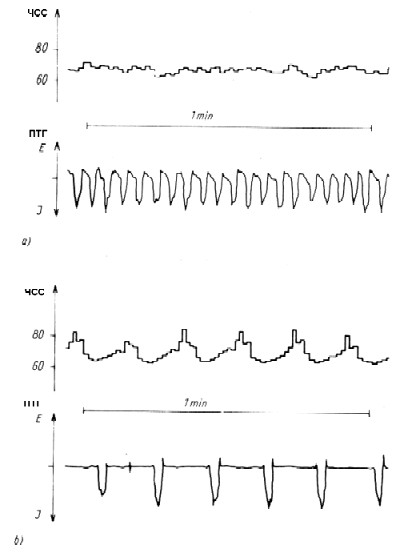
Рис.24. Изменение крутизны характеристических кривых регуляции дыхания по СО2 после практики пранаям (Stanescu и др. 1981).








Какую роль, таким образом, могут играть различные пранаямы в системе регуляции внешнего дыхания? Прежде всего, надо уяснить себе в принципе, что произвольные манипуляции формой дыхания с целью получить определенные изменения минутных объемов дыхания все же не составляют непосредственной цели пранаямы. Что касается глубины и частоты дыхания, то они находятся в обратных соотношениях: быстрые пранаямы выполняются с относительно меньшим объемом вдоха, чем медленные, а там, где на первом плане стоит глубина дыхания, падает его частота («великое дыхание», уджайи, мурча). При всех пранаямах с углубленным вдохом (по аналогии с процессами в скелетной моторике при асанах) до предела своих функциональных возможностей должны быть задействованы проприорецепторы, что ведет к аналогичным эффектам, а именно к усилению афферентации с увеличением специфической и неспецифической репрезентации в ЦНС (см.3.6). В условиях покоя это означает также «сознательное» противодействие рефлексу Геринга-Брейера. Во всех случаях экстремально замедленного дыхания (кумбхака, уджайи, мурча, бхрамари, кевали, а также ритмическое дыхание) следовало бы ожидать гипоксию и гиперкапнию, но, к сожалению, до сих пор не проводилось соответствующих измерений парциальных давлений газов крови. Медленные пранаямы зачастую выполняются очень долго, в экспериментах по самопогребению до 9 часов (см.5.3.2). Такие практики, по-видимому, ведут также и к адаптивным процессам химической регуляции дыхания. При исполнении и вследствие уджайи меняется крутизна характеристических кривых регуляции дыхания по CO2 (Stanescu и др. 1981), что равным образом означает снижение активности системы химической регуляции дыхания, а стало быть, в перспективе становление нечувствительности регулятора. На рис.24 показано различие средних кривых регуляции дыхания по CO2 для восьми испытуемых с многолетним опытом хатха-йоги (асаны, уджайи и кумбхака) и кривых регуляции по CO2 восьми членов контрольной группы. При дыхательном тестировании в замкнутом объеме было обнаружено при сравнении спонтанного дыхания и уджайи (рис.25), что ответная зависимость дыхания от напряжения CO2 при уджайи понижается.
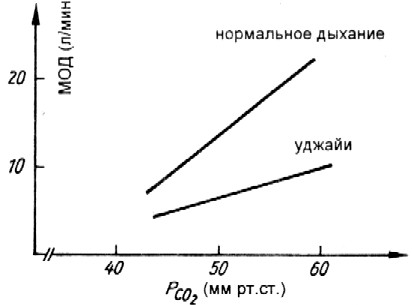
Рис.25. Сравнение спонтанного дыхания и уджайи при дыхании в замкнутом объеме (при уджайи ответное увеличение вентиляции на рост парциального давления СО2 несколько ниже; Stanescu с соавт. 1981).
Здесь находит свое выражение непосредственное воздействие формы дыхания уджайи (сознательно измененной) на величину дыхания, т.о. «преодолевается» химическая регуляция дыхания. К сожалению, это единственное исследование в своем роде, и было бы показательно исследовать также воздействие других пранаям. Bhole (1967, см.5.3.2) сообщает, что йоги учили подавлять непроизвольную реакцию химической регуляции дыхания на раздражение углекислым газом. Это указание следует также и из экспериментов по самопогребению медитирующих йогов (см.5.3.2), в которых хотя и было установлено увеличение в выдыхаемом воздухе углекислого газа, но не замерялась ни форма дыхания, ни даже парциальные давления дыхательных газов в крови. У последовательных йогов, которые соблюдают также и рекомендации по диете (см.2.3), можно было бы предположить «изменение должных значений» химической регуляции дыхания вследствие манипуляций с условиями метаболизма, как например, ощелачивание крови, изменение буфрных систем из-за сдвигов в йонном фонде и т.п. Хотя исследований на эту тему не проводилось. Неизвестно, каких размеров при таких практиках достигает гипоксия. Впрочем, при кратковременном исполнении уджайи вновь было обнаружено повышение потребления кислорода (Miles 1964, Ebert, Waurik 1980).
Существенная проблема пранаямы, которую вообще-то не принято обсуждать в физиологической литературе, - характер психического влияния на форму дыхания. С одной стороны, все упражнения пранаямы выполняются произвольно в том смысле, что они реализуются лишь при желании практикующего - в иных случаях он дышит спонтанно. С другой стороны, при исполнении пранаям требуется некоторая психическая дистанция к процессу дыхания в смысле пассивной концентрации (см.5.1). Следует по возможности без эмоций наблюдать как «вообще дышится» и глубину дыхания изменять потому, что «оно» дышит глубже, но не потому, что «я» глубже дышу. Это выглядит так, как если бы процесс дыхания полностью передавался центру автоматизма и форма дыхания менялась бы не произвольно, но как если бы занимающийся лишь наблюдал за тем, как она меняется. Для того, чтобы дать этому приемлимое объяснение, необходимо сперва, до выдвижения каких-бы то ни было психофизиологических гипотез, психологически более точно сформулировать понятия «я», «оно» (не в психоаналитическом смысле), «произвольно», «сознательно» и т.д.
Физиологические процессы, коррелирующие с дыханием
Наряду с тем, что дыхание подвержено многочисленным воздействиям, дыхание также оказывает влияние на множество иных функций организма.
При этом не всегда возможно точно определить, являются ли эти функциональные связи целенаправленными или случайными, соответственно, имеют ли они смысл или нет. Различные периодические процессы могут взаимодействовать так, что одна из функций доминирует, а значит ее ритм накладывается на ритм другой функции (суперпозиция), или функции входят во временные соотношения, а значит, каждая функция изменяет свою частоту таким образом, что возникают целочисленные соотношения частот. Von Holst (1939) назвал устойчивое целочисленное соотношение частот абсолютной координацией, при которой сохраняется постоянное соотношение фаз, тогда как соотношение, наступающее только с некоторой статистической вероятностью, - относительной координацией. Von Holst описал феномены этих координаций на примерах локомоторных ритмов насекомых, рыб и млекопитающих. Относительно процессов дыхания на сегодня известны следующие взаимосвязи с координацией ритмов.
Дыхательный ритм находится в целочисленном частотном отношении к спонтанному ритму ходьбы (Anders 1928), сосательному ритму младенцев (Peiper 1938), к ритму спортивных движений (Михайлов и др. 1971), к ритму бега на велоэргометре (Hildebrandt, Daumann 1965), к произвольным движениям глаз (Waurik 1981), а у свободнолетающих птиц - к ритму взмахов крыльев (Lord и др. 1962, Berger и др. 1970). Эти частотные соотношения показывают, что дыхательная моторика включена в общую моторику и зависит от нее. По мнению von Holst"а смысл такого рода взаимосвязей ритмических процессов состоит в оптимизации энергетического обмена: затраты энергии меньше, если два процесса, протекающих взаимосвязанно, имеют постоянные соотношения фаз при целочисленных соотношениях частот. То же вытекает из математических моделей отношений между взаимосвязанными осцилляторами (Dörre 1977).
Из вариантов наложения ритмов наиболее известны амплитудная модуляция кровяного давления с частотой дыхания и модуляция частоты сердечных сокращений (дыхательная аритмия). Кроме того, существуют еще обусловленные дыханием ритмические вазомоторные сокращения (во время вдоха) мышц (Koepchen 1968) и слизистой носа (Eccles, Lee 1981). Такого рода влияния дыхательного ритма на вегетативные функции могут проходить двумя способами: