
3.4 Изменчивость шишек. Сравнение.








Ель аянская: зрелые шишки изящные, коричневые, длиной 3-8 см, толщиной 1,5-3 см, цилиндрически удлиненные, мягкокожистые, рыхлые из-за волнистых семенных чешуй, немного изогнутые, созревают в ноябре.
Ель сибирская: шишки маленькие, тем более в неблагоприятных лесорастительных условиях. Семенные их чешуи широкие и закругленные. Женские шишки после опыления красноватые, прямостоячие, при созревании темно-коричневые, висячие, яйцевидно-овальные, 3,5-8 см длины, 2,5 см в диаметре.
3.5 Изменчивость коры. Сравнение.
Ель аянская: ствол старых деревьев с серо-бурой, растрескивающейся и отслаивающейся небольшими пластинками корой.
Ель сибирская: кора темно-серая или почти черная, морщинистая.
3.6 Изменчивость побегов. Сравнение.
Ель аянская: побеги блестящие, желто-бурые или буро-серые, с сильно отстоящими листовыми подушками, изогнутыми у основания, и небольшим сегментовидным возвышением.
Ель сибирская: побеги красновато-коричневые до желто-серых или почти белых, часто опушены короткими, редкими, рыжеватыми волосками, реже почти голые.
3.7 Изменчивость почек. Сравнение.
Ель аянская: почки желто-бурые до бледно-золотистых, яйцевидно-конические, всегда несмолистые, плотно покрыты приостренными чешуйками с изорванными верхушками.
Ель сибирская: почки от яйцевидных до овальных, красно-коричневые, несмолистые, почечные чешуйки плотно прилегающие, из них нижние часто имеют шиловидное острие.
3.8 Изменчивость семенных чешуек. Сравнение.
Ель аянская: семенные чешуи кожистые, мягкие, продолговато-овальные, продольно-желобчатые, с выемкой наверху, с ясно зазубренными краями, снаружи беловатые или светло-коричневые, блестящие, продольно-слабо-струйчатые.
Ель сибирская: семенные чешуи тонкие, выпуклые, вееровидные, на верхнем конце округлые, реже отсеченные, слегка притупленные, цельнокрайние, иногда слабо выемчатые. Чешуйка коричнево-желтая, блестящая, слегка покрыта матовым коротким пушком.
3.9 Изменчивость кроющих чешуек. Сравнение.
Ель аянская: кроющие чешуи скрыты, в 3-4 раза меньше семенных, эллиптические, коротко заостренные, продолговатые.
Ель сибирская: кроющие чешуи в 5 раз короче семенных, на верхушке грубозубчатые, как бы выгрызенные.
3.9 Изменчивость семян. Сравнение.
Ель аянская: семена мелкие (2 мм), коричневые или почти черные, в 2 раза мельче семян ели обыкновенной (до 5 мм длины и 3 мм ширины). Крылышко по краю мелко зазубрено.
Ель сибирская: семена мелкие, черноватые, яйцевидные до бледно-желтоватых, с крылом в 3 раза длиннее семени.
4 Методы изучения наследственности
4.1 Мутагенез
Мутагенез — процесс возникновения наследственных изменений организма — мутации. Редкое появление среди нормальных организмов измененных особей было известно давно. Однако научное описание явлений мутагенез было сделано лишь в 1899 г. рус. ученым С. И. Коржинским и в 1900—1901 гг. гол. генетиком Г. де Фризом, в частности, ввел термины «мутация» (лат. изменение) и «мутагенез».
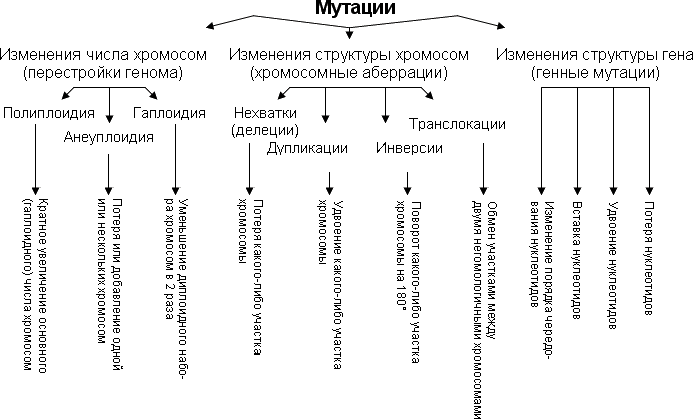 Рисунок 8. Виды мутаций.
Рисунок 8. Виды мутаций.
Виды мутаций
Существует несколько классификаций мутаций по различным критериям. В современной учебной литературе используется и более формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций:
· геномные
· хромосомные
· генные
Геномные — полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосом) и анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору [Инге-Вечтомов, 1989].
В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай — объединение целых хромосом, т. н.
Робертсоновская транслокация, которая является переходным вариантом от хромосомной мутации к геномной).
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях.
Цитоплазматические мутации – изменения в ДНК митохондрий и пластид. Передаются только по женской линии, т.к. митохондрии и пластиды из сперматозоидов в зиготу не попадают. Пример у растений – пестролистность.
Соматические – мутации в соматических клетках (клетках тела; могут быть четырех вышеназванных видов). При половом размножении по наследству не передаются. Передаются при вегетативном размножении у растений, при почковании и фрагментации у кишечнополостных (у гидры).
Мутагенные факторы применительно к лесным древесным породам делят на физические и химические. К основным физическим мутагенам относятся: радиация, температурные шоки, ультрафиолетовые лучи. В мутационной селекции наиболее широко применяются ионизирующие излучения, которые по своей природе подразделяются на волновые и корпускулярные. К волновым излучениям относятся ультрафиолетовые лучи, рентгеновские лучи и гамма-лучи. К корпускулярным излучениям относятся а-частицы, протоны, нейтроны, дейтроны и др. Ионизирующие излучения при проникновении в клетку действуют непосредственно на наследственные структуры [Корнеев, 1985].
Наименьшей проникающей способностью обладают ультрафиолетовые лучи, рентгеновские и гамма-лучи. Проникающая способность корпускулярных излучений выше, чем у гамма-лучей, более чем в 20 раз. Источниками ультрафиолетовых лучей, эффективных при облучении пыльцы, являются ртутно-кварцевые лампы, рентгеновских лучей— рентгеновские установки, гамма-лучей—радиоактивные изотопы ( Са и др.), корпускулярных излучений — ядерные колонки [Корнеев, 1985].
Химические мутагены вызывают преимущественно генные мутации, физические — генные и хромосомные. Характер мутаций изучают на основе оценки чувствительности растения к мутагену. Оказалось, что слабое воздействие мутагена стимулирует процессы жизнедеятельности растений. С усилением воздействия мутагеном стимулирующий эффект возрастает и достигает предела, затем постепенно падает и снижается до нуля. После этого он оказывает противоположное действие — начинает подавлять процессы жизнедеятельности и тем больше, чем сильнее воздействие. Подавляющее воздействие мугагенов возрастает с повышением дозы воздействия до полной гибели клеток, органа или всего растения.
Различают стимулирующие, критические, летальные и оптимальные дозы мутагенов. Дозы мутагена определяются мощностью источника излучения, концентрацией и продолжительностью их воздействия на растения. Дозы, при которых всхожесть семян составляет около 50 % контроля, а выживаемость 20—30% числа всходов, называются критическими. Дозы, вызывающие гибель обрабатываемого материала, называются летальными, а дозы, при которых на единицу выживаемости растения получается наибольшее количество мутаций, называются оптимальными.
Чувствительность растений к мутагенам можно оценивать по энергии прорастания семян, всхожести, выживаемости растений, степени плодовитости и стерильности, энергии роста, числу и типу хромосомных перестроек в первом митозе в клетках проростков. По чувствительности к мутагенам древесные растения делят на три группы:
1) чувствительные—кедр сибирский, лиственница, сосна обыкновенная, ели, дуб черешчатый, березы, ольха, смородина черная, шелковица (диплоидная), яблони (грушовка, белый налив) и др.
2) среднечувствительные—рябина обыкновенная, шиповник, акация желтая, сирень, жасмин, боярышник, жимолость обыкновенная, орех черный, каштан конский, шелковица (тетраплоидная), облепиха (европейского происхождения), яблони (антоновка, пепин шафранный) и др.
3) устойчивые—липа, тополь черный, гледичия, спирея, ракитник, облепиха (алтайская) [Любавская, 1982].
Предложенный мной способ получения мутанта.
Данный способ требует наименьших затрат труда. Он наиболее пригоден для работы с культурами, которые имеют быстро прорастающие семена. Обработке можно подвергать как сухие, так и предварительно замоченные в воде семена. По окончании обработки, перед посевом их хорошо промывают в проточной воде. Прежде всего, перед тем как сажать обработанные семена, нужно провести культивацию земли. Затем Сухие семена замачить в растворе колхицина (0,1—0,2%) в течение 3—6 дней, и после чего начинать сеять. Иногда семена можно погрузить в воду до набухания, а затем прорастить в течение 0,5—48 ч в чашках, на фильтровальной бумаге, смоченной раствором колхицина.
4.2 Полиплоидия
Полиплоидия - это увеличение числа наборов хромосом в клетках организма, кратное гаплоидному (одинарному) числу хромосом; тип геномной мутации. Половые клетки большинства организмов гаплоидны (содержат один набор хромосом – n), соматические – диплоидны (2n). Организмы, клетки которых содержат более двух наборов хромосом, называются полиплоидами: три набора – триплоид (3n), четыре – тетраплоид (4n) и т. д. Наиболее часто встречаются организмы с числом хромосомных наборов, кратным двум, – тетраплоиды, гексаплоиды (6 n) и т. д. Полиплоиды с нечётным числом наборов хромосом (триплоиды, пентаплоиды и т. д.) обычно не дают потомства (стерильны), т. к. образуемые ими половые клетки содержат неполный набор хромосом – не кратный гаплоидному [Инге-Вечтомов, 1989].
Полиплоидия может возникнуть при нерасхождении хромосом в мейозе. В этом случае половая клетка получает полный (нередуцированный) набор хромосом соматической клетки (2n). При слиянии такой гаметы с нормальной (n) образуется триплоидная зигота (3n), из которой развивается триплоид. Если обе гаметы несут по диплоидному набору, возникает тетраплоид. Полиплоидные клетки могут возникнуть в организме при незавершённом митозе: после удвоения хромосом деления клетки может не происходить, и в ней оказываются два набора хромосом. У растений тетраплоидные клетки могут дать начало тетраплоидным побегам, цветки которых будут вырабатывать диплоидные гаметы вместо гаплоидных. При самоопылении может возникнуть тетраплоид, при опылении нормальной гаметой – триплоид. При вегетативном размножении растений сохраняется плоидность исходного органа или ткани.
Полиплоидия широко распространена в природе, но среди разных групп организмов представлена неравномерно. Большое значение этот тип мутаций имел в эволюции диких и культурных цветковых растений, среди которых ок. 47 % видов – полиплоиды [Котов, 1997г.].
Для получения полиплоида, можно использовать такой способ как «опыление нередуцированными гаметами».
Данные гаметы получают путём нагревания пыльцы в термостате при температуре 38-40 (градусов Цельсия), в течении 1,5-3 часов, с последующим её выдерживанием при температуре 18-20 (градусов Цельсия), и влажности воздуха 70-80 (процентов). Такой пыльцой опыляют опытное растение. Условия для опытных растений должны быть оптимальными (влажность, температура). Отбор полиплоида проводят сначала по морфологическим признакам, затем путём цитологического анализа. Всё морфологические изменения определяются по контрольному растению (растение не подверженное человеческим вмешательством).
4.3 Генная инженерия
Генная или генетическая инженерия – это совокупность биотехнологическихметодов, позволяющих создавать синтетические системы на молекулярно-биологическом уровне.
Генная инженерия дает возможность конструировать функционально активные структуры в форме рекомбинантных нуклеиновых кислот: рекДНК (recDNA) или рекРНК (recRNA) – вне биологических систем (in vitro), а затем вводить их в клетки.
Задачи генной инженерии
Основные направления генетической модификации организмов:
– придание устойчивости к ядохимикатам (например, к определенным гербицидам);
– придание устойчивости к вредителям и болезням (например, Bt-модификация);
– повышение продуктивности (например, быстрый рост трансгенного лосося);
– придание особых качеств (например, изменение химического состава).
Методы генной инженерии
Методы генной инженерии основаны на получении фрагментов исходной ДНК и их модификации.
Для получения исходных фрагментов ДНК разных организмов используется несколько способов:
– Получение фрагментов ДНК из природного материала путем разрезания исходной ДНК с помощью специфических нуклеаз (рестриктаз).
– Прямой химический синтез ДНК, например, для создания зондов.
– Синтез комплементарной ДНК (кДНК) на матрице мРНК с использованием фермента обратной транскриптазы (ревертазы).
В состав вектора ДНК входит не менее трех групп генов:
1. Целевые гены, которые интересуют экспериментатора.
2. Гены, отвечающие за репликацию вектора, его интеграцию в ДНК клетки-хозяина и экспрессию требуемых генов.
3. Гены-маркеры (селективные, репортерные гены), по деятельности которых можно судить об успешности трансформации (например, гены устойчивости к антибиотикам или гены, отвечающие за синтез белков, светящихся в ультрафиолетовом свете).
Практические достижения современной генной инженерии заключаются в следующем:
– Созданы банки генов, или клонотеки, представляющие собой коллекции клонов бактерий. Каждый из этих клонов содержит фрагменты ДНК определенного организма (дрозофилы, человека и других).
– На основе трансформированных штаммов вирусов, бактерий и дрожжей осуществляется промышленное производство инсулина, интерферона, гормональных препаратов. На стадии испытаний находится производство белков, позволяющих сохранить свертываемость крови при гемофилии, и других лекарственных препаратов.
– Созданы трансгенные высшие организмы (многие растения, некоторые рыбы и млекопитающие) в клетках которых успешно функционируют гены совершенно других организмов. Широко известны генетически защищенные генно-модифицированные растения(ГМР), устойчивые к высоких дозам определенных гербицидов, а также Bt-модифицированные растения, устойчивые к вредителям. Среди трансгенных растений лидирующие позиции занимают: соя, кукуруза, хлопок, рапс [Лобашев, 1979г.].
Заключение
Темой для написания моей курсовой работы стала ель аянская. Так как древесина используется наравне с древесиной ели европейской, но по механическим свойствам уступает ей. Имеет декоративность, ее придают синевато-белые полоски на нижней стороне хвои, благодаря ее изогнутости крона кажется сизой. Перспективна для создания контрастных групп, выделяющихся голубовато-сизой хвоей на фоне темнохвойных пород.
Для человека ель — одно из самых полезных деревьев. Очень широко используется еловая древесина. Именно из нее изготавливают лучшие сорта бумаги, искусственный шелк, шерсть и многое другое, используют в строительстве. Особенно важна древесина ели в изготовлении музыкальных инструментов (скрипок, альтов, контрабасов, пианино). Именно ее использовали знаменитые мастера Страдивариус, Амати, Гварнери. Для музыкальных инструментов в лесах отбирают 100 — 120-летние деревья с особой древесиной, у которой годичные кольца одинаковой толщины. Такие ели получили название «резонансовых». Отобранные для изготовления инструментов ели подсекали и на 3 года оставляли на корню. При этом дерево постепенно теряло влагу и древесина уплотнялась, становилась более легкой, благодаря чему инструменты получали особую силу звучания [Бобров, 1971г.].
В работе была изучена изменчивость, наследственность и генная инженерия ели аянской. Предложен свой способ получения мутанта и полиплоида.
Библиографический список
1. Алёшин, Е. П./ Алёшин, Н. Е.- Рис. Москва, 1993. 504 стр. 100
2. Бобров Е.Г. История и систематика рода Picea A. Dietr.
// Новости систематики высших растений. Л., 1971. Вып. 7. С. 5-40.
3. Булыгин, Н. Е. Дендрология / Н. Е. Булыгин [и др.]. - Москва: МГУЛ,
2010.
4. Вавилов, Н. И. Теоретические основы селекции / Н. И. Вавилов – Москва:
Наука, 1987.
5. Инге-Вечтомов, С.Г. Генетика с основами селекции / С. Г. Инге-Вечтомов. -
Москва: Высшая школа, 1989.
6. Колесников, А. И. Дендрология / А. И. Колесников. – Москва: Лесная
промышленность, 1969.
7. Котов, М. М. Генетика и селекция / М. М. Котов. - Йошкар-Ола: МарГТУ,
1997.
8. Лобашев М.Е., Ватти К.В., Тихомирова М.М. Генетика с основами селекции: Учеб. пособие для студентов пед. ин-тов по биол. спец. – М.: Просвещение, 1979. – 304 с.
9. Любавская, А. Я. Лесная селекция и генетика / А. Я. Любавская. - Москва:
Лесная промышленность, 1982.
10. Любавская, А. Я. Лесная селекция и генетика / А. Я. Любавская. - Москва:
МГУЛ, 2007
11. Мамаев С.А. Основные принципы методики исследования внутривидовой изменчивости древесных растений //
Индивидуальная эколого-географическая изменчивость растений. Свердловск, 1975. Вып. 94. С. 3–14.
12. Муратова Е. Н. 1994. Хромосомный полиморфизм в при- родных популяциях лиственницы Гмелина Larix gmelinii (Rupr.) Rupr. Цитология и генетика. 28(4) : 14—22.
13. Муратова Е. Н. 1995а. Методики окрашивания ядрышек для кариологического анализа хвойных. Бот. журн. 80(2) : 82—86.
14. Муратова Е. Н. 19956. Кариосистематика семейства Pi- naceae Lindl Сибири и Дальнего Востока: Автореф. докт. дис. Новосибирск. 32 с. Муратова Е. Н. 2000. В-хромосомы голосеменных. Успе- хи соврем, биол. 120(5): 452—465.
15. Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Советская наука, 1952. 390 с.
16. Серебряков И.Г. О морфогенезе жизненной формы дерева у лесных пород средней полосы Европейской части СССР // Бюлл. МОИП. Отд. биологии. 1954. Т. LIX(1). С. 53-68.
17. Серебряков И.Г. Экологическая морфология растений: Жизненные формы покрытосеменных и хвойных. М., 1962. 378 с.
18. Царёв, А. П. Генетика лесных древесных растений / А. П. Царёв [ и др.]. –
Москва: МГУЛ, 2010.