
2 физиологические индикаторы внимания








Как известно, внимание характеризуется определенными физиологическими показателями.
Разбирая работу первого функционального блока головного мозга — блока, обеспечивающего общий тонус коры, — мы указывали на тот факт, что всякое явление активации (arousal) сопровождается целым комплексом симптомов. Сюда относятся хорошо известные изменения сердечной деятельности и дыхания, сужение периферических сосудов, кожно-гальваническая реакция, явление десинхронизации (или депрессии альфа-ритма) и др.
Кэтим хорошо известным коррелятам внимания за последние годы присоединились и другие показатели изменения функциональных состояний: медленные потенциалы, получившие название «волн ожидания» (expectancy waves), введенные Греем Уолтером, увеличение числа синхронно работающих пунктов коры (М.Н.Ливанов, 1962, 1967), изменения среднего уровня асимметрии восходящего и нисходящего фронтов альфа-волн, описанные А.А.Генкиным (1962, 1963, 1964; и др.) и др.
263
Все эти явления носят выраженный генерализованный характер, и их можно считать признаками изменения функционального состояния или общей внимательности субъекта.
Наряду с этим существуют, однако, и другие ЭЭГ-явления, позволяющие объективно выявить специализированные формы направленного, избирательного внимания.
К таким явлениям относятся изменения параметров вызванных потенциалов коры головного мозга.
Вызванные потенциалы, впервые описанные Эдрианом (1936), а затем многими другими авторами — Жувэ (1957), Эрнандес-Пеоном (1961, 1966), Даусоном (1958, 1959), И.А.Пеймером (1957) и др., — как известно, представляют собой электрические ответы на различные (зрительные, слуховые и др.) раздражители в соответствующих (затылочных, височных и др.) областях мозговой коры и подкорковых структурах. Параметры вызванных потенциалов изменяются в зависимости от интенсивности раздражителя, с одной стороны, и активности субъекта — с другой, причем изменения параметров вызванных потенциалов могут распространяться на ранние фазы ответа (т.е. возникать в интервале 15 — 100 мс) или касаться более поздних фаз (от 100 мс до 250 — 300 мс и больше).
Параметры вызванных потенциалов могут быть использованы не только как индикатор относительно элементарной анализаторной деятельности мозга, они с успехом могут быть использованы и для объективной регистрации тех изменений в приеме и переработке информации, которые возникают при мобилизации активного внимания.
Изучение вызванных потенциалов как объективного индикатора внимания можно использовать двумя путями.
С одной стороны, можно изучать изменения вызванных потенциалов при отвлечении внимания побочным раздражителем, с другой стороны, — в условиях привлечения внимания к данному раздражителю. Первый вариант опыта был проведен в классических исследованиях Эрнандес-Пеона (1956, 1960, 1969), показавшего, что корковые звуковые потенциалы резко тормозятся при предъявлении кошке вида или запаха мыши. Второй вариант опыта был проведен преимущественно на людях и заключался в том, что вызванные потенциалы в ответ на сенсорные раздражители сравнивались с вызванными потенциалами, полученными в условиях активного ожидания этих сигналов (соответственно предупреждающей инструкции) или в условиях усложнения анализа этих раздражителей и т. п. Факты, полученные в этих условиях рядом исследователей (Линдсли, 1960, 1961; А.И.Пеймер, 1958, 1966; Э. Г. Симерницкая, 1970; Тесце, 1970; и др.), показали, что привлечение внимания активным ожиданием или усложнением задачи приводит к заметному повышению амплитуды поздних компонентов вызванных потенциалов по сравнению с «фоном». Таким образом, изменение параметров потенциалов (особенно поздних компонентов) является отчетливым объективным признаком селективного внимания.
264
Как показали многочисленные исследования, введение задания (например, переход к инструкции внимательно следить за изменениями сигнала) приводит не только к увеличению амплитуды вызванных потенциалов, но и к распространению на другие зоны коры головного мозга, лежащие вне пределов коркового «ядра» данного анализатора.
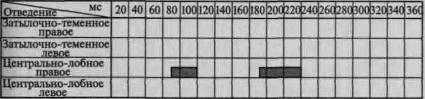
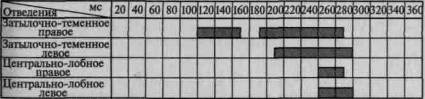
Пример такого вовлечения вторичных и третичных отделов коры по мере мобилизации внимания соответствующей инструкцией в вызванную активность можно видеть на рис. 101. Этот рисунок отчетливо показывает, что предварительная инструкция не только увеличивает амплитуду вызванного потенциала, но и ведет к его
1. Ожидание тока
2. Дифференцирование зрительных стимулов
3.Выполнение произвольных двигательных реакций
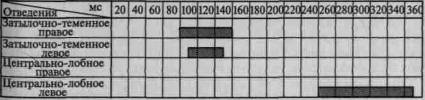
Рис. 101. Распространение достоверных изменений вызванных
потенциалов на отдельные зоны коры при активации избирательного
внимания (по Э. Г. Симерницкой)
распространению на другие более широкие области коры. Данные изменения параметров вызванных ответов при привлечении внимания взяты из исследования, проведенного в нашей лаборатории Э. Г. Симерницкой (1970).
Объективные электрофизиологические исследования позволяют не только изучать физиологические механизмы различных видов внимания; они дают также возможность проследить основные этапы их формирования. Эту возможность показали работы, проведенные Д. А. Фарбер и ее сотрудниками (Д.А. Фарбер, 1969; Е. М. Фрид, 1970; Д.А.Фарбер, Е.М.Фрид, 1971). Установлено, что если рефлекс вызывает заметное повышение амплитуды вызванных потенциалов даже у маленького ребенка, то стойкие изменения вызванных потенциалов в результате речевой инструкции (считать сигналы или наблюдать за их изменениями) носят у него еще недостаточно выраженный и устойчивый характер и начинают отчетливо и стойко выступать лишь на позднейших этапах развития ребенка.
266
Эти данные, приведенные на рисунках 102 и 103, показывают, что физиологические изменения, вызываемые речевой инструкцией и лежащие в основе произвольного внимания, формируются лишь к 12—15-летнему возрасту. Один из наиболее существенных для нас фактов заключается в том, что именно к этому возрасту выраженные и стойкие изменения в вызванных потенциалах начинают возникать не только в сенсорной области коры, но и в ее лобных отделах, и что лобные отделы коры, по всем данным, начинают к этому времени принимать ближайшее участие в обеспечении сложных и устойчивых форм высшего, произвольного внимания.
К специальному рассмотрению этого вопроса, представляющего большой нейропсихологический интерес, мы и обратимся.
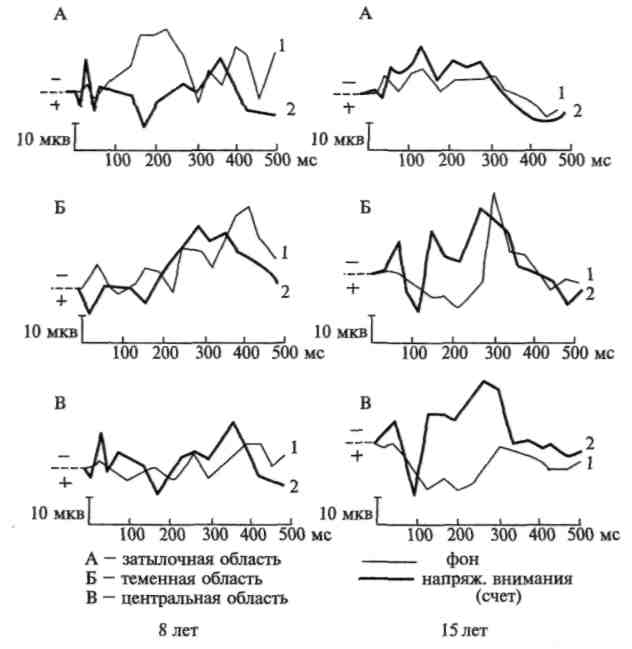
Рис. 102. Увеличение амплитуды вызванных потенциалов при напряжении активного внимания (по Д. А. Фарбер и Е. М. Фрид)
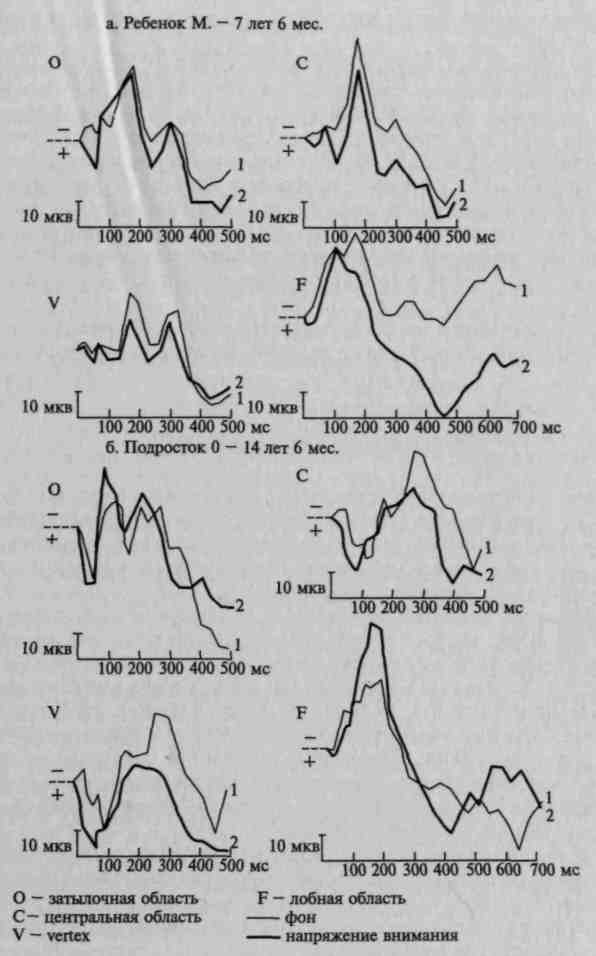
Рис. 103. Распространение изменений в вызванных потенциалах на
различные зоны коры при мобилизации активного внимания у детей
разного возраста. Изменение вызванных потенциалов под влиянием
речевой инструкции оценивать различные зрительные сигналы:
а — у ребенка 7 лет 6 мес; 6 — у подростка 14 лет 6 мес. (по Д. А. Фарбер и Е. М. Фрид)
3 МОЗГОВАЯ ОРГАНИЗАЦИЯ
Есть все основания считать, что не все системы переднего мозга играют одинаковую роль в процессах организации внимания и что вопрос о мозговой организации внимания следует дифференцировать как в отношении отдельных структур мозга, так и в отношении отдельных форм внимания.
Рассматривая вопрос об основных функциональных блоках головного мозга, мы уже указывали ранее на ту роль, которую играют образования верхних отделов ствола и ретикулярной формации среднего м о з г а в поддержании уровня бодрствования и в появлении общей реакции пробуждения (arousal). Именно поэтому после работ Мэгуна и Моруцци (1949), Линдсли и др. (1950), Джаспера (1956) данные образования считались основными механизмами, определяющими переход от сна к бодрствованию и обеспечивающими наиболее генерализованные и элементарные формы внимания.
Эти предположения подтвердились не только большим числом широко известных опытов с животными, при которых перерезка стволовой ретикулярной формации на определенном уровне приводила ко сну, а ее раздражение — к повышению бодрствования и обострению чувствительности (см. обзор этих фактов Линдсли, 1960), но и рядом клинических наблюдений, установивших, что поражения верхних отделов ствола и стенок третьего желудочка могут приводить к возникновению сна или к онейроидному, про-соночному состоянию и что тонус коры в этих случаях оказывается резко сниженным, а селективные формы сознания и внимания — глубоко нарушенными.
268
Однако стволовые и мезэнцефалические механизмы восходящей активирующей ретикулярной формации обеспечивают лишь наиболее генерализованные состояния бодрствования и внимания.
Для обеспечения избирательных (селективных) форм внимания, т. е. выделения того или другого сигнала и торможения реакций на побочные раздражители, необходимо участие более высоко расположенных образований головного мозга неспецифических ядер таламуса — лимбической коры и лобной области больших полушарий.
За последнее время накоплено много данных, освещающих ту роль, которую играют образования древней коры, или лимбической области (гиппокамп, миндалина), и связанные с ними аппараты хвостатого ядра в регуляции внимания и бодрствования. Исследования, проведенные на нейронном уровне, показали, что именно в этих образованиях (особенно в гиппокампе) значительное место занимают нейроны, отвечающие не на модально-специфические раздражения, а производящие как бы сличение (компарацию) старых и новых раздражителей и обеспечивающие реакцию на новые сигналы или их свойства с угашением реакции на старые, уже привычные раздражители (О.С.Виноградова, 1969, 1970). Поэтому образования гиппокампа, интимно участвующие в обеспечении механизмов торможения посторонних раздражителей и привыкании (habituatiori) к длительно повторяющимся раздражителям, стали считать одной из существенных составных частей того тормозящего или «фильтрующего» аппарата, который необходим для избирательных реакций на специфические раздражители, входящие в систему врожденных ориентировочных рефлексов и инстинктивного поведения. Именно в силу этого гиппокамп, а затем и хвостатое ядро стали рассматривать как существенные аппараты, обеспечивающие организму возможность осуществлять строго избирательные формы поведения (Граштиан, 1961; Дуглас, Прибрам, 1966; О.С.Виноградова, 1969, 1970), а нарушение этих образований — как источник распада селективности поведения в целом, которое в большей степени можно отнести к нарушениям избирательного внимания, чем к дефектам памяти. Этим представлениям о роли лимбических структур, и в частности образований гиппокампа, в организации поведения отвечают и те изменения в поведении и в состоянии сознания, которые наблюдаются у человека при поражениях этой области, а также при изучении в специальных электрофизиологических исследованиях.
Клинически больные этой группы — с расположенными по средней линии глубокими опухолями — не дают отчетливых нарушений гнозиса и праксиса, речи или формально-логических процессов. Патология психических процессов у данных больных заключается в выраженном нарушении избирательности, селективности психических процессов, в резкой их истощаемости.
269
В относительно более стертых случаях это проявляется в повышенной отвлекаемости больных, быстром прекращении активной, направленной деятельности, в легком всплывании побочных ассоциаций и в отчетливо выступающих нарушениях памяти, к которым мы еще обратимся далее. В более тяжелых случаях (массивные опухоли, влияющие на стенки третьего желудочка и лимбические структуры) этот синдром может принимать грубо выраженные формы и переходить в просоночные онейроидные состояния, при которых больные теряют грань между настоящим и прошлым, обнаруживают явления спутанности, конфабуляции. В обоих случаях любые посторонние раздражители сразу же вплетаются в течение мыслей, и организованное, избирательное строение сознания уступает место спутанности. Такие больные могут заявить, что они находятся не в больнице, а дома или на работе (а иногда и там, и здесь), что тут же рядом находятся их родные, они принимают врача за знакомого по месту работы и дают признаки грубых расстройств сознания, которые оказываются особенно отчетливо выраженными, если в патологический процесс вовлекаются передние отделы лимбической области и если критика к собственному состоянию нарушается (А. Р. Лурия, Е. Д.Хомская, С.М.Блинков, М.Кричли, 1967).
Описанные клинические факты нашли свое частичное разъяснение в тех электрофизиологических данных, которые были получены в последнее время при исследовании больных с массивными поражениями диэнцефальной области, опухолями стенок третьего желудочка и примыкающих к ним отделов лимбической системы.
В этих исследованиях (Л.П.Латаш, 1968; Е.Д.Хомская, 1969, 1972; Н.А.Филиппычева, Т.О.Фаллер, 1970; и др.) было отчетливо показано, что вегетативные и электрофизиологические компоненты ориентировочного рефлекса в этих случаях оказываются резко угнетенными, и даже повторные раздражители начинают вызывать лишь очень слабые и быстро угасающие реакции, что является отчетливым результатом дефицита тех неспецифических активирующих влияний, которые в норме обеспечивают соответствующий тонус коры мозга.
Закон силы, по которому сильные раздражители вызывают сильные, а слабые раздражители — слабые реакции, нарушается, разные по интенсивности раздражения начинают вызывать одинаковые слабые реакции, часто предъявленное раздражение вызывает не депрессию, а парадоксальную экзальтацию альфа-ритма. Процесс привыкания к новому раздражителю (habituation), отчетливо наблюдаемый при нормальном состоянии коры, также претерпевает грубые изменения, и электрофизиологические показатели ориентировочного рефлекса либо вообще не вызываются, либо становятся неугасимыми.
270
Одновременно эти исследования показали, что у данной группы больных можно наблюдать и известные извращения нормального протекания нервных процессов, что вплотную подводит нас к объяснению механизмов только что описанных психических явлений.
Существенным для этих случаев оказывается тот факт, что все эти признаки первичного нарушения селективного внимания, по данным ЭЭГ, могут в той или иной степени компенсироваться введением речевой инструкции, иначе говоря, включением в систему более сохранных высших уровней построения процесса. Чаще всего эта компенсация имеет лишь временный характер, и избирательность процессов активации быстро теряется; однако эта потенциальная сохранность высших, произвольных форм внимания при первичном нарушении его элементарных форм является одним из важных признаков, отличающих этих больных от больных с иной локализацией поражения (Е.Д.Хомская, 1972).
Совершенно иную роль в организации внимания играют лобные доли головного мозга.
В классических исследованиях над животными, как уже упоминалось ранее, считалось, что лобные доли мозга имеют непосредственное отношение к сохранению следов памяти, что и проявляется в факте нарушения отсроченных реакций после экстирпации лобной коры. Однако, как известно, этот факт получил в дальнейших работах совершенно иное истолкование (Малмо, 1942; Прибрам, 1959, 1963; Вейзкранц, 1968; и др.). Показано, что животное с удаленными лобными долями не могло сохранить отсроченных реакций не столько потому, что оно не удерживало прежних следов, а вследствие постоянного отвлечения побочными раздражителями. Устранение этих побочных раздражителей (например, путем помещения животного в полную темноту или проведение опытов на фоне транквилизаторов, снижающих общий тонус коры) приводило к восстановлению отсроченных реакций. Существенная роль лобных долей в торможении реакции на любые побочные раздражители и в сохранении направленного, программированного поведения была показана и опытами Конорского (1964; и др.), Брутковского (1964, 1966) и др. Именно в силу нарушения тормозящих механизмов экстирпация лобных долей неизменно приводила у животных к грубым нарушениям целенаправленного, избирательного поведения и к расторможению реакций на побочные раздражения. Соответствующие факты изменения поведения животных описаны в классических работах Бианки (1895, 1921), Джекобсена (1935), П. К.Анохина (1949), Прибрама (1954, 1958; и др.) и др.
271
Богатейшие данные о нарушениях избирательного поведения, и прежде всего высших произвольных форм внимания, дает клиника поражений лобных долей мозга.
В отличие от описанных случаев поражений верхних отделов ствола и лимбической области элементарные формы непроизвольного внимания или импульсивно возникающие ориентировочные реакции на любые раздражители могут быть у таких больных не только сохранены, но даже и патологически усилены; наоборот, всякие попытки вызвать у этих больных устойчивое произвольное внимание по речевой инструкции не приводят ни к каким результатам.
Невозможность сосредоточиться на данной инструкции и затормозить реакции на любые побочные раздражители становятся видны уже при обычных клинических наблюдениях над больными с массивными поражениями лобных долей мозга. Обычно такие больные начинают выполнять предложенное им задание, но лишь только в палату входит посторонний человек или сосед по палате обращается к кому-либо шепотом, больной тотчас же перестает выполнять начатое задание, переводит взор на вошедшего в палату или вмешивается в разговор соседа. Именно поэтому, чтобы исследовать такого больного, надо проводить опыт с его соседом: в этих случаях он непроизвольно вплетается в начатый разговор, обнаруживая при этом ту непроизвольную активность, которую трудно вызвать, обращаясь к нему с непосредственно адресованными ему заданиями.
Эта повышенная отвлекаемость больного с массивным поражением лобных долей становится источником глубоких нарушений его целенаправленного поведения.
Глубокое нарушение произвольного внимания или высших форм контроля над деятельностью при оживлении элементарных форм внимания и создает ту картину нарушений поведения, которую мы так часто встречаем в клинике поражений лобных долей мозга.
Ряд психофизиологических исследований, проведенных за последние годы, позволяет подойти к выделению механизмов, лежащих в основе нарушения высших произвольных форм внимания.
Как уже указывалось ранее (ч. вторая, гл. V), Грей Уолтер установил факт, что именно в лобных долях человека ожидание какого-либо сигнала вызывает специфические медленные потенциалы, названные им «волнами ожидания», и что лишь впоследствии эти волны распространяются на другие участки мозга (рис. 48). По данным М.Н.Ливанова и его сотрудников, каждое интеллектуальное напряжение вызывает в лобных отделах коры закономерное повышение числа синхронно работающих точек (рис. 49).
272
Эти явления указывают на непосредственное участие лобных долей мозга человека в активации, вызываемой речевой инструкцией; лобные доли являются частью мозговых систем, которые непосредственно вовлекаются в процессы, связанные с высшими формами активного внимания. Наличие богатых нисходящих связей лобных долей мозга с лимбической системой и ретикулярной формацией ствола является фактом, позволяющим выделить и некоторые морфофизиологические основы участия лобных долей в этих высших формах активации.
При психофизиологических исследованиях больных с поражением лобных долей мозга многолетними и тщательными исследованиями Е. Д.Хомской и ее сотрудников был установлен ряд фундаментальных фактов (Е.Д.Хомская, 1960, 1965, 1966, 1969, 1971, 1972; Е.Д.Хомская, А.Р.Лурия, 1970; Е.Д.Артемьева, 1965, 1966; О.П.Барановская, 1966, 1968; Э.Г.Симерницкая, 1966, 1970; и др.).
Как уже упоминалось, массивные поражения верхних отделов ствола среднего диэнцефального мозга и лимбической области могут привести к нарушениям первичных основ внимания — ориентировочного рефлекса, который либо угнетается и легко угасает, либо, напротив, не подчиняется факторам привыкания. Характерно, однако, что в этих случаях привлечение внимания к сигналу путем речевой инструкции может в известных пределах компенсировать этот дефект и усиливать и стабилизировать электрофизиологические и вегетативные компоненты ориентировочного рефлекса. Аналогичное можно наблюдать и у больных с поражениями задних отделов мозга, у которых патологическое состояние коры может делать физиологическую основу внимания неустойчивой, но у которых, однако, речевая инструкция, данная больному, сразу же повышает тонус коры и в определенной мере компенсирует дефект. Настойчивую работу над компенсацией дефекта можно наблюдать у этих больных и в процессе восстановительного обучения, она, безусловно, требует напряженности и стойкости произвольного внимания.
Совершенно иное можно наблюдать у больных с массивными поражениями лобных долей мозга.
Как уже говорилось, элементарные ориентировочные рефлексы у этих больных часто оказываются патологически усиленными, и именно это мешает им целенаправленно выполнять задания.
Другая картина обнаруживается у этих больных, если наблюдать их реакции на последовательно предъявляемые сигналы. Значительная часть больных этой группы обнаруживает в этих случаях большую нестойкость внимания, и уже через несколько предъявлений одних и тех же раздражителей вегетативные и электрофизиологические компоненты ориентировочного рефлекса исчезают.
273
Отличие больных с поражениями лобных долей от больных с очаговыми поражениями в задних отделах мозга заключается, однако, в другом: если у последних больных привлечение произвольного внимания дополнительной инструкцией (например, инструкцией «считать сигналы», «следить за их изменением» и т.п.) сразу же восстанавливает и стабилизирует угасшие компоненты ориентировочного рефлекса, то у больных с поражением лобных долей мозга этого не происходит. Инструкция, которая должна повысить активность больного, не приводит ни к каким сколько-нибудь устойчивым сдвигам в системе ориентировочных реакций.
Это проявляется в целой серии показателей процессов активации.
Так, если в норме многократное повторение сигнала приводит к угашению сосудистой ориентировочной реакции, достаточно предложить испытуемому следить за изменениями интенсивности или длительности сигнала или считать количество сигналов, сосудистые реакции, являющиеся компонентом ориентировочного рефлекса, сразу же восстанавливаются и остаются устойчивыми в течение длительного времени. Этого нет у больных с массивными поражениями лобных долей мозга; сосудистые реакции на повторяющиеся сигналы у них быстро угасают, и речевая инструкция, в норме мобилизующая внимание, не приводит у них ни к какому эффекту: кривая плестизмограммы остается неизменной.
Далее, в норме первые предъявления сигналов вызывают отчетливые сдвиги в электроэнцефалограмме в виде депрессии альфа-ритма и повышения высоких частот спектра; многократное повторение сигналов приводит к явлениям привыкания, т.е. к исчезновению этих реакций: после инструкции, привлекающей внимание к этим сигналам, ЭЭГ-компоненты ориентировочного рефлекса восстанавливаются вновь и приобретают устойчивый характер. Эта стабилизующая и активизирующая функция внимания, сохраняющаяся у больных с поражениями задних отделов больших полушарий, нарушается у больных с массивными поражениями лобных долей мозга, и если даже они и в состоянии выполнять данную им инструкцию, то это протекает без явной мобилизации внимания по ЭЭГ-показателям.
Наконец, как известно, в норме раздражение определенной модальности приводит к появлению вызванных потенциалов в соответствующих отделах мозговой коры; введение инструкции различать сигналы приводит к заметному усилению амплитуды поздних компонентов вызванных потенциалов и их большему распространению по коре головного мозга. Эти явления, подробно описанные рядом авторов, не наблюдались у больных с массивными поражениями лобных долей; хотя у них, как и у других испытуемых, сигнал вызывал отчетливый вызванный потенциал, параметры этого потенциала не менялись под влиянием инструкции, мобилизующей внимание.
274
Соответствующие иллюстрации приводились на рисунке 86.
Все эти данные дают полное основание предполагать, что лобные доли мозга принимают ближайшее участие в повышении уровня бодрствования соответственно поставленным перед субъектом задачам и тем самым играют решающую роль в обеспечении высших произвольных форм внимания. Этот факт, несомненно имеющий большое значение для понимания мозговых механизмов сложных форм внимания, был детально изучен в работах Е.Д.Хомской и ее сотрудников, и сравнительные наблюдения над различными формами поражения лобных долей мозга позволили не только установить только что сформулированное положение, но и существенно уточнить его.
Оказалось, что описанное явление нарушения регуляции процессов активации посредством речевой инструкции проявлялось особенно отчетливо у больных с массивными двусторонними поражениями лобных долей мозга, захватывающими ее медиальные или медиобазальные области. Это выступало как у тех больных, которые проявляли в своем поведении уже описанные выше черты инактивности, отвлекаемости и глубокого нарушения избирательности психических процессов, так и у больных, которые клинически не проявляли заметных для внешнего наблюдения симптомов изменения поведения.
Больные с поражениями конвекситальных отделов лобных долей мозга обнаружили другую картину. После угашения ориентировочных реакций, возникавших у них под влиянием сигналов, вызвать их усиление и стабилизацию с помощью речевой инструкции оказывалось все же возможным, хотя лишь после многократного повторения инструкции. Однако в этих случаях они оказывались нестойкими и быстро исчезали.
Все эти исследования имеют большое значение для установления мозговых механизмов высших произвольных форм внимания. Они показывают, что лобные доли мозга играют важную роль в организации произвольной сознательной деятельности человека.
Глава IV ПАМЯТЬ
ПСИХОЛОГИЧЕСКОЕ СТРОЕНИЕ
Изучение механизмов памяти долгое время было малоразработанным разделом нейрофизиологии; только за последнее десятилетие появились работы в этом направлении. Мозговая организация различных форм мнестической деятельности остается в значительной степени новой главой нейрофизиологической науки.
Причиной такого положения в большой степени являются упрощенные взгляды на память, которых придерживается подавляющее большинство физиологов, и недооценка сложности структуры мнестических процессов, продемонстрированной за последние 20 — 30 лет (здесь и далее в подобных случаях иметь в виду, что первое издание книги вышло в 1973 году. — примеч. ред.) психологами.
Если резюмировать все, что было известно в психологии о природе и о материальном субстрате памяти к началу этого века, становится ясно, насколько бедна была информация, которой располагала наука.
С одной стороны, она включала положение Р.Семона и Э.Геринга о том, что память, или «способность сохранять следы», является «всеобщим свойством материи», — положение, которому нельзя отказать в правильности, но которое имеет слишком общий характер. С другой стороны, бытовали представления А. Бергсона (1896) о том, что существуют два вида памяти: «память тела» и «память духа» и что если первая относится к числу естественных явлений — тех самых, о которых говорили Семон и Геринг, — то вторая должна рассматриваться как проявление «свободной воли», способной путем духовного, волевого усилия вызывать индивидуальные следы прежнего опыта.
Сравнительно мало обогатили вопрос о природе памяти и морфофизиологические исследования первых четырех десятилетий этого века. Тщательное морфологическое исследование нервной клетки и ее связей не пошло дальше общего утверждения, что сохранение следов раз возникших возбуждений является, по-видимому, результатом наличия синаптического аппарата (Рамон-и-Кахал, 1909—1911) и что в основе его, по-видимому, лежат некоторые биохимические процессы, связанные с равновесием
276
ацетил-холина и холин-эстеразы, играющих основную роль в синаптической передаче импульса (де Робертис, 1964; Экклз, 1957— 1966). Работы по физиологии условного рефлекса, целиком посвященные процессам выработки и закрепления опыта, установили лишь некоторые основные физиологические факторы того, что весьма условно обозначалось как «проторение путей» или «закрепление условных связей», но фактически не сделали каких-либо существенных шагов для раскрытия природы памяти. Не достигли больших успехов и исследователи процесса научения (learning) — в основном американские бихевиористы. Несмотря на то что число публикаций, посвященных этому вопросу, исчислялось многими тысячами, вопрос о природе памяти оставался совершенно неясным. Именно это и дало основание выдающемуся американскому психологу К. С.Лешли в его известной статье «В поисках энграммы» (1950) прийти к пессимистическому заключению, что материальная природа памяти остается такой же нераскрытой, какой она была многие десятилетия назад.
Существенный сдвиг в понимании материальных основ памяти был сделан в результате работ Хидена (I960, 1962, 1964), показавшего, что сохранение следа от возбуждения связано с изменением структуры рибонуклеиновой кислоты, и проследившего закономерное возрастание содержания РНК—ДНК в ядрах, подвергавшихся интенсивному возбуждению. Работы Хидена положили начало целому потоку исследований, и положение, что молекулы РНК—ДНК являются носителями памяти, что именно они играют решающую роль как в наследственной передаче, так и в прижизненном сохранении следов памяти, вскоре стало общепризнанным.
Из этого положения был даже сделан вывод, что информация, полученная одной особью, может передаваться гуморальным путем второй особи; получившие широкую огласку (но остающиеся дискуссионными) опыты Мак-Коннелла и Джекобсона (1970) вселяли надежду на то, что материальные основы памяти будут вскоре окончательно раскрыты.
К этим исследованиям присоединились другие, которые показали участие в сохранении следов раз возникшего возбуждения не только нервной клетки, но и окружающей ее глии. Эти исследования показали, что процессы возбуждения в нейроне и в глии (в моменты запечатления) протекают с разным латентным периодом (который в глии в несколько сот раз больше, чем в нейроне) и находятся в реципрокных отношениях, выражающихся в том, что в момент возбуждения уровень РНК в нейроне повышается, а в окружающей глии падает, в то время как в последующем периоде (видимо, связанном с сохранением следа) он резко падает в нейроне, но столь же резко и длительно возрастает в глии. Положение об участии глии в процессах сохранения следов, бесспорно, относится к числу наиболее важных открытий современной нейрофизиологии и, по-видимому, раскрывает некоторые интимные механизмы нейрофизиологии памяти.
277
Важные данные также были получены в исследованиях последних лет, проводившихся с помощью электронной микроскопии и связанных с прямыми наблюдениями над нервной клеткой в состоянии возбуждения. Установлены факт движения мельчайших пузырьков (везикул) в процессе возбуждения и факт изменения мембран в процессе образования следов (де Робертис, 1964; Экклз, 1957, 1961), что также, видимо, имеет непосредственное отношение к интимным механизмам следообразования.
Исследования механизмов следовых процессов на клеточном и субклеточном (молекулярном) уровнях существенно продвинули изучение интимных биохимических и морфофизиологических механизмов памяти. Однако они не дали прямых ответов на вопрос, какие зоны мозга реально участвуют в процессах памяти и какие стороны сложных форм мнестической деятельности обеспечиваются теми или другими системами мозга.
Для того чтобы приблизиться к ответу на этот вопрос, необходимо было, с одной стороны, от общих и слишком диффузных представлений о следовых процессах перейти к четким психологическим представлениям о реальной структуре мнестической деятельности. С другой стороны, нужно было перейти от исследований следовых процессов на клеточном и молекулярном уровнях к изучению реальной мозговой архитектоники процессов памяти, иначе говоря, к анализу того, что именно вносит каждая из зон мозга в организацию мнестических процессов человека.
Эти вопросы являются предметом постоянного внимания психологов, с одной стороны, и неврологов и нейропсихологов — с другой, и за последние двадцать лет наши представления о строении мнестических процессов обогатились благодаря целой серии оригинальных исследований.
Попытаемся резюмировать их в самом кратком виде, с тем чтобы затем перейти к основному содержанию этой главы — анализу того, какую роль в организации мнестических процессов играют те или другие аппараты головного мозга человека.
Классическая психология трактовала запоминание либо как процесс непосредственного запечатления («записи») следов в нашем сознании, либо как процесс запечатления тех однозначных ассоциативных связей, в которые вступают друг с другом отдельные впечатления. Это упрощенное представление о процессе запоминания оказалось несостоятельным.
278
Как показал ряд работ, опубликованных за последнее время (Норман, 1966, 1968, 1970; Викельгрен, 1970; Кинч, 1970; Куби, 1969; Познер, 1963, 1967, 1969; Шифрин, 1970; Рейтман, 1970), запоминание представляет собой сложный, развернутый во времени, процесс, который распадается на ряд последовательных этапов, характеризующихся неодинаковой психологической структурой, неодинаковым «объемом» доступных для фиксации следов и неодинаковой длительностью их хранения.
Ряд авторов (Сперлинг, 1960, 1963, 1970; Мортон, 1969, 1970) высказали предположение, что процесс запоминания начинается с запечатления различных сенсорных признаков (например, фонетических признаков услышанного слова). В момент запечатления выделяются только некоторые из этих признаков, т. е. производится определенный отбор (Норман и др., 1970; Викельгрен, 1970; Сперлинг, 1970; и др.). Некоторые авторы характеризуют эту стадию как ультракороткую память, так как этап запечатления узок по объему и очень короток по длительности запечатлеваемых следов (Бродбент, 1970).
Этот этап мнестического процесса многие понимают как перевод сигналов на уровень иконической памяти, при этом воспринятые раздражители превращаются в кратковременные образы (images). За этим этапом следует процесс образования кратковременной памяти, который предполагает выбор соответствующего образа из многих возможных и может быть интерпретирован как своеобразная переработка или кодирование полученных сигналов (Кинч, 1970; Шифрин, 1970; Рейтман, 1970; Познер, 1969).
Этот этап рассматривается, однако, большинством авторов лишь как промежуточный, уступающий место последнему этапу, заключающемуся в сложном кодировании следов, или включении их в некоторую систему категорий.
Тщательный анализ категориальных связей, в которые включается след каждого запечатлеваемого субъектом воздействия (или получаемой им информации), является центральным предметом большого числа исследований мнестических процессов, опубликованных за последнее время. Некоторые из них носят чисто психологический или логико-гипотетический характер (Норман и др., 1968, 1970; Викельгрен, 1970; Кинч, 1970; Познер, 1963, 1969), другие представляют собой попытки построить сложные модели памяти, исходящие из представлений о роли такого кодирования (Рейтман, 1970; Фейгенбаум, 1970), третьи исходят в своем анализе из психолингвистических концепций (Дж. Миллер, 1969; Мортон, 1969, 1970). Все они, однако, единодушно приходят к утверждению, что системы связей, в которые вводятся следы доходящей до субъекта информации, строятся на основе различных кодов и, следовательно, представляют собой многомерные системы, из которых субъект должен каждый раз производить выбор соответствующей системы. Такой подход к процессам памяти полностью отходит от понятий, согласно которым запоминание пред-
279
ставляет собой однозначный и пассивный процесс, и подчеркивает сложную и активную природу процессов запоминания. Этот подход обращает внимание на тот факт, что человек, запоминающий известный материал, обнаруживает известную стратегию запоминания, выбирая нужные средства, выделяя существенные и тормозя несущественные признаки, выделяя соответственно задаче то сенсорные, то смысловые компоненты запечатлеваемого материала и компонуя его в соответствующие системы (Кинч, 1970; Шифрин, 1970; Познер, 1963, 1969; Рейтман, 1970).
Не меньшее внимание, чем проблема запоминания, привлекла к себе и тесно связанная с нею проблема забывания. Какие причины лежат в основе трудностей актуализации нужных следов? Еще поколение назад ответ на этот вопрос казался относительно простым. Исследователи, начиная с Эббингауза (1885), молчаливо принимали тот факт, что со временем каждый след, оставленный тем или иным раздражителем, стирается; действительно, проверка, проведенная через несколько часов или дней после первоначального запечатления следа, дает возможность наблюдать естественное забывание его.
В последние десятилетия предположение, что забывание является естественным следствием постепенного угасания следов (trace decay), было высказано рядом авторов (Браун, 1958, 1964; Конрад, 1960). Эта гипотеза вызвала, однако, ряд возражений. Толчком к ним были следующие факты. Прежде всего, с течением времени можно наблюдать иногда не угасание, а, наоборот, повышенное воспроизведение следов, которое получило в психологии специальное название реминисценции. Далее, фактом, не согласующимся с теорией, предполагающей, что угасание следов является механизмом забывания, было наличие ряда ошибочных воспроизведений специфического характера, которые часто наблюдались у испытуемых по истечении некоторого периода времени. Наконец, фактом, который не согласуется с описанным пониманием забывания, было отрицательное влияние на процесс воспроизведения всякой побочной деятельности, отделяющей момент запечатления от момента воспроизведения.
Прежние представления о забывании как о пассивном процессе требовалось заменить новыми. Еще в начале XX века Мюллер и Пильцеккер (1900) выступили с предположением о том, что забывание является скорее результатом тормозящего влияния со стороны побочных, интерферирующих воздействий, чем следствием постепенного угасания следов. К этому предположению присоединились и другие авторитетные исследователи (Робинсон, 1920; Скэггс, 1925; Мак-Фи, 1932; Мелтон, 1940, 1941; А. А. Смирнов, 1941; Андервуд, 1957, 1960, 1966; Постман, 1961, 1963, 1967; Кеппель, 1968), внимательно изучавшие тормозящее влияние на следы памяти как предшествующих, так и последующих воздей-
280
ствий. В настоящее время положение о том, что явления «проактивного» и «ретроактивного» торможения следует рассматривать как очень существенные факторы забывания, прочно вошло в литературу, и господствующей теорией мнестических процессов стала теория, согласно которой забывание является в значительной мере следствием влияния побочных, интерферирующих воздействий, тормозящих нормальное воспроизведение ранее запечатленных следов.
В настоящее время подтвердились представления о структуре мнестических процессов, сформулированные отечественными психологами еще в 30-х годах (Л. С. Выготский, А. Н.Леонтьев и др.). Вкратце они сводились к тому, что память лишь в относительно редких случаях является у человека элементарной, непосредственной и что, как правило, процесс запоминания опирается на систему вспомогательных средств и, таким образом, является опосредованным. Для объективного изучения процесса опосредования (или, как мы бы теперь сказали, кодирования) материала, подлежащего запоминанию, был предложен ряд специальных приемов (А. Н.Леонтьев, 1931; и др.).
Одновременно другие исследователи (А.А.Смирнов, 1947, 1966) дали подробный анализ роли активной смысловой организации в процессе запоминания материала и показали богатство различных приемов, которые применяются при запоминании осмысленного материала взрослым человеком. В специальных исследованиях отечественных психологов (П. И. Зинченко, 1961), подвергавших детальному изучению процесс непроизвольного запоминания, и была выяснена та роль, которую играет в нем стоящая перед субъектом задача (детерминация направления внимания и отбора запоминаемого материала).
Все эти исследования позволили убедиться в том, что запоминание представляет собой сложный активный процесс, или, иначе говоря, специальную форму сложной и активной мнестической деятельности. Эта мнестическая деятельность определяется задачей запомнить соответствующий материал; для ее осуществления требуются определенная стратегия и соответствующие средства и коды, увеличивающие объем запоминаемого материала, длительность его удержания, а иногда, как это было отмечено в специальных исследованиях, приемы, снимающие тормозящее действие побочных интерферирующих агентов, которое, как мы уже говорили, лежит в основе забывания.
Все эти исследования, описывающие сложную психологическую структуру мнестических процессов человека, открывают пути для изучения мозговых механизмов памяти как мнестической деятельности.
281
Совершенно естественно, что процесс строго направленного, избирательного запоминания предполагает оптимальный тонус коры, или состояние ее полного бодрствования. Далее процесс активного запоминания предполагает наличие у субъекта намерения запомнить, при отсутствии или недостаточной стойкости которого эффективность процесса не может быть обеспечена. Кроме того, описанный выше сложный и распадающийся на ряд последовательных ступеней процесс приема и кодирования информации предполагает также полную сохранность корковых отделов соответствующих анализаторов, которые должны быть в состоянии разложить поступающую информацию на ряд дробных, модально-специфических (зрительных, слуховых, тактильных и т.д.) признаков, осуществить процесс отбора этих признаков и, наконец, перейти к объединению их в целые динамические структуры.
Наконец, переход от элементарного (сенсорного) этапа получения и запечатления информации к более сложным этапам организации ее в образы и затем к кодированию ее в категориально-организованные системы требует сохранности наиболее сложно-организованных вторичных и третичных зон коры, одни из которых связаны с синтезом последовательно поступающих сигналов в сукцессивные, или симультанные, структуры, а другие — с организацией этих следов в языковые коды.
Все это говорит о том, что сложный по своей структуре процесс запоминания опирается у человека на целую систему совместно работающих аппаратов мозговой коры и нижележащих образований и что каждый из этих аппаратов вносит свой специфический вклад в организацию мнестических процессов.
Поэтому есть основания ожидать, что разрушение или даже патологическое состояние любого из этих аппаратов неизбежно скажется на протекании мнестических процессов и что нарушения мнестической деятельности будут иметь неодинаковый характер в зависимости от того, какое именно звено окажется пострадавшим.
МОДАЛЬНО-НЕСПЕЦИФИЧЕСКИЕ ФОРМЫ ПАМЯТИ
Как уже было сказано ранее, основным условием запечатления любых следов является сохранение оптимального тонуса коры. Снижение коркового тонуса является основным фактором, который делает невозможным избирательное запечатление и хранение следов и приводит к нарушению общей модально-неспецифической памяти. Поэтому очевидно, что изучение физиологических механизмов обеспечения условий сохранения оптимального тонуса коры имеет непосредственное отношение к исследованию физиологических основ общей способности запечатлевать и сохранять избирательные системы следов, иначе говоря, к исследованию мозговых механизмов элементарной, общей памяти.
282
Как известно, первые шаги в этом направлении были сделаны В.М.Бехтеревым (1900), который впервые высказал предположение, что поражения медиальных отделов височной области могут привести к нарушениям памяти, и Грюнталем (1939), который показал, что при поражении мамиллярных тел, являющихся релейными ядрами для волокон, идущих от гиппокампа в составе круга Пейпеца, возникают тяжелые расстройства памяти.
Значительно позднее в работах Сковилла (1954), Сковилла и Милнер (1957, 1967), Пенфилда и Милнер (1958) и Милнер (1958, 1962, 1966, 1968, 1969, 1970) было показано, что двусторонние поражения гиппокампа не затрудняют протекания высших познавательных процессов, но существенно нарушают общую способность запечатлевать следы текущего опыта и приводят к нарушениям памяти, близким к классическому корсаковскому синдрому.
Эти данные получили двоякое морфофизиологическое объяснение. С одной стороны, многочисленные исследования, посвященные морфологическому и физиологическому анализу стволовых и таламических неспецифических образований и их путей показали, что лимбические отделы мозга, и в частности гиппокамп, являются образованиями, обеспечивающими модуляцию тонуса коры, и что поражение этих отделов старой коры неизбежно приводит к снижению тонуса коры, а отсюда и к нарушению возможности избирательного запечатления следов.
С другой стороны, позднейшие исследования на нейронном уровне показали, что именно в гиппокампе и связанных с ним лимбических структурах имеется значительное число нейронов, не реагирующих на какие-либо модально-специфические раздражители, но обеспечивающих компарацию актуальных раздражений со следами прошлого опыта; эти нейроны реагируют на каждое изменение раздражений и, таким образом, являются в одинаковой мере нейронами внимания и нейронами памяти (рис. 30).
Эти данные дают основания предполагать, что, изучая функции ретикулярной формации, неспецифических ядер таламуса и образований круга Пейпеца, мы действительно приближаемся к мозговым аппаратам, обеспечивающим общую функцию запечатления следов, которая является первой и наиболее элементарной основой памяти.
Анализ клинического материала полностью подтверждает это предположение.
Как уже говорилось в четвертой главе, участие в мнестических процессах медиальных отделов коры головного мозга (в частности гиппокампа) и связанных с ними образований легко доказывается теми нарушениями памяти, которые возникают при их поражениях. Больные даже с относительно нерезко выраженными признаками дисфункции этих отделов мозга, возникающими, напри-
283
мер, при опухолях гипофиза, вторично влияющих на образования лимбического мозга, проявляют отчетливые нарушения памяти, которые не только выступают в жалобах больных, но могут быть обнаружены также путем объективного исследования. У больных с массивными поражениями медиальных отделов мозга, захватывающими образования круга Пейпеца (двусторонние поражения гиппокампа, мамиллярных тел и т.д.), эти нарушения принимают грубейший характер и приводят к полной невозможности сохранять следы текущего опыта, нередко проявляясь в синдроме грубой дезориентированности больного в месте, времени и происходящих с ним событиях.
Три свойства характерны для этих нарушений памяти.
Во-первых, они имеют модально-неспецифический характер, так что их можно выявить в любой сфере деятельности больного (забывание намерений, забывание только что выполненных действий, впечатлений и т. п.).
Во-вторых, они одинаково проявляются как в элементарном, непреднамеренном запечатлении следов, так и в специальной произвольной мнестической деятельности. Это с полной отчетливостью демонстрируется тем фактом, что некоторые из больных со стертыми формами нарушения памяти могут даже обращаться к ряду вспомогательных приемов, например к записыванию намерений с целью компенсирования своих дефектов. Эта особенность характерна для больных с двусторонним поражением гиппокампа, описанных Б. Милнер (1958—1966).
Наконец, в-третьих, в случаях массивных поражений указанных структур мозга нарушения памяти сопровождаются грубыми нарушениями сознания, чего никогда не наблюдается у больных с локальными поражениями конвекситальных отделов мозговой коры. Как уже было сказано, в наиболее грубых случаях эти больные теряют представление о том, где они находятся, не могут дать правильную оценку времени, правильно ориентироваться в окружающем, т. е. проявляют те симптомы растерянности и спутанности, которые часто встречаются в психиатрической клинике, но которые относительно редко можно наблюдать в клинике локальных поражений мозга.
Описанные признаки отчетливо показывают, что поражения глубоких отделов мозга и медиальных отделов коры приводят к первичным расстройствам памяти, которые никак не связаны с какими-либо частными дефектами познавательной (аналитико-синтетической) деятельности, и что, следовательно, глубокие отделы мозга, расположенные по средней линии (ретикулярная формация ствола, таламуса, лимбические образования), имеют прямое отношение не только к поддержанию оптимального тонуса коры, но и к обеспечению условий, необходимых для сохранения непосредственных следов.
284
Экспериментальные исследования, которые были проведены в нашей лаборатории за последние годы (А.Р.Лурия и др., 1970; А.Р.Лурия, А.Н.Коновалов, А.Я.Подгорная, 1970; Л.Т.Попова, 1964; Н.К.Киященко, 1969, 1973; А.Р.Лурия, 1973), позволили более тщательно охарактеризовать первичные расстройства памяти, возникающие в результате глубоких поражений мозга, и наметить их физиологические механизмы.
Не повторяя того, что уже было изложено ранее, остановимся на самых основных из полученных нами результатов.
Первые попытки найти основные проявления первичных нарушений памяти при описываемых поражениях не привели к обнадеживающим результатам. Как показали наблюдения, больные даже с массивными нарушениями памяти, приближающимися к корсаковскому синдрому, нередко оказывались в состоянии воспроизвести серию из 5—6 прочитанных ими слов, не отличаясь существенно от здоровых испытуемых. Этот факт заставлял думать, что дефект, характеризующий нарушение памяти в интересующих нас случаях, не лежит в самом процессе запечатления (записи) поступающей (пусть в относительно ограниченных пределах) информации. Дальнейшие опыты показали, что короткие ряды элементов могут удерживаться этими больными в течение небольшого срока (1—2 мин, иногда больше) только тогда, когда этот промежуток времени остается незаполненным какой-либо другой, посторонней, деятельностью. Это означало, что основой наблюдаемого дефекта, по-видимому, не является слабость следов. Факты исчезновения следов по истечении более длительного промежутка времени, наблюдавшиеся у этих больных, оставались неубедительными из-за того, что невозможно было установить, действовали на испытуемого в течение этих промежутков посторонние раздражители или нет.
Важные данные были получены в опытах, в которых экспериментатор ставил перед собой задачу проследить, в какой степени изучаемые следы изменяются под влиянием побочных отвлекающих воздействий. Проведение их было вызвано, с одной стороны, тем фактом, что, как правило, переданная больному информация или только что произведенное действие тотчас же исчезали из его памяти, как только внимание его отвлекалось посторонним раздражителем или другой деятельностью, а с другой — требовалось проверить разделяемую рядом авторов (Мелтон, 1943, 1960, 1970; Андервуд, 1945, 1957, 1960; Постман, 1954, 1969) теорию, согласно которой раз возникшие следы сохраняются на длительный срок, а в основе заболевания лежит не столько угасание следов (trace decay), сколько торможение следов побочными (интерферирующими) воздействиями.
285
Для того чтобы проверить гипотезу, согласно которой патологическое состояние мозга, возникающее вследствие нарушения нормальных активирующих неспецифических влияний на кору головного мозга, приводит к патологической тормозимости возникших следов, была проведена серия опытов (А. Р.Лурия, 1971, 1973; А.Р.Лурия, А. Н. Коновалов, А.Я.Подгорная, Н. К.Киященко, 1969, 1973). Первый из этих опытов заключался в том, что наряду с проверкой удержания элементов данной серии (слов, фраз, картинок, действий) после паузы, не заполненной никакой побочной деятельностью («пустая пауза»), проводился опыт, отличающийся от описанной ситуации тем, что пауза заполнялась какой-либо посторонней деятельностью (например, счетом), после чего испытуемый должен был снова воспроизвести уже воспроизведенную ранее серию.
Второй опыт сводился к тому, что испытуемому предлагалось сначала запомнить одну (очень короткую) серию, состоявшую из двух-трех слов, фраз или картинок, после чего ему давалась для запоминания вторая аналогичная серия, а вслед за этим от него требовалось вспомнить, какую серию он запоминал в первый раз.
Эти серии опытов дали следующие результаты.
У нормального испытуемого переход к выполнению другой, побочной, деятельности существенно не отражался на припоминании первого ряда, а запоминание второго, короткого, ряда элементов не мешало воспроизведению ранее запоминавшегося первого ряда. У больных с поражениями глубоких отделов мозга и синдромом общего (первичного) нарушения памяти, легко удерживавших серию из трех-четырех элементов без отвлечения, после побочной (интерферирующей) деятельности (например, счета) возможность припомнить ранее удержанный ряд либо полностью исчезала, либо больной воспроизводил лишь изолированные фрагменты ряда. Например, если после запоминания группы из трех-четырех слов больному предлагалось запомнить вторую аналогичную группу, возвращение к первой оказывалось полностью невозможным, и ее следы, казалось, полностью стирались.
Близкие явления, отчетливо говорящие о повышенной тормозимости следов побочными интерферирующими воздействиями, отмечались не только при припоминании серии слов, но и при всякой другой мнестической деятельности (запоминании серий картинок, движений, фраз и рассказов). Таким образом, патологически повышенная тормозимость следов носила общий, модально-неспецифический характер.
Наиболее отчетливые результаты были получены в опытах с припоминанием первой серии слов (фраз, картинок, действий) после запоминания второй такой же серии. Ретроактивное торможение, вызванное второй группой слов, оказывалось в этих случаях столь большим, что первая группа следов либо полностью исчезала из памяти больного, либо при воспроизведения больной смешивал следы первой и второй группы, т. е. контаминировал их.
286
И в этом случае повышенная тормозимость следов и утрата их избирательности проявлялись независимо от модальности и, что самое важное, не снимались фактором организации следов в осмысленные структуры.
Так, запомнив сначала фразу «В саду за высоким забором росли яблони», а затем вторую — «На опушке леса охотник убил волка», больной с выраженным модально-неспецифическим нарушением памяти либо совсем не мог припомнить первую фразу, либо же смешивал (контаминировал) ее элементы с элементами второй фразы и воспроизводил что-то вроде: «В саду... на опушке охотник убил волка» или «На опушке леса... росли яблони». То же обнаруживалось и при воспроизведении первого рассказа после прочтения второго: содержание первого рассказа часто оказывалось полностью забытым.
Тормозимость следов интерферирующими воздействиями у наиболее тяжелых больных оказывалась настолько резко выраженной, что выполнение даже какого-либо простого действия нацело забывалось после того, как больной выполнял второе такое же действие: больной, нарисовавший некоторую фигуру, не только забывал ее после рисования второй фигуры, но даже не соглашался с тем, что показанная ему первая фигура была действительно нарисована им самим.
Характерно, что многократное повторение опыта обычно не приводит к улучшению результатов, и этот факт показывает, что патологически повышенное взаимное торможение следов является основным физиологическим фактором первичных нарушений памяти, наблюдаемых при глубоких поражениях мозга.
Приведенные данные позволяют сделать выводы об основных физиологических механизмах интересующих нас первичных, модально-неспецифических нарушениях памяти.
Не меньший интерес представляют также факты, дающие возможность судить об уровне таких нарушений. Как мы уже указывали выше, относительно стертые нарушения функций медиальных отделов коры (возникающие, например, при опухолях гипофиза, выходящих за пределы турецкого седла и влияющих на образования лимбической области) проявляются в повышенной тормозимрсти следов, которая выступает лишь на уровне удержания серий изолированных следов (слов, картинок) и проявляется особенно отчетливо при действии интерферирующих раздражений. Удержание организованных серий следов (фраз, рассказов), а также более элементарных сенсомоторных следов остается здесь сохранным и не поддается тормозящему влиянию интерферирующих воздействий.
287
Другой характер нарушений можно наблюдать в случаях массивных поражений глубоких отделов мозга (расположенных по средней линии), приводящих к грубым расстройствам памяти.
В этих случаях первичные мнестические расстройства распространяются как на более элементарные уровни, так и на более высокоорганизованные структуры следов, и нарушения памяти приобретают более генеральный характер.
Если вызвать у нормального испытуемого (или у больного со стертыми формами нарушения неспецифической памяти) фиксированную гаптическую установку методом, предложенным Д.Н.Узнадзе, — когда испытуемому несколько раз предлагается ощупывать правой рукой больший, а левой меньший шар, после чего левый из двух одинаковых (контрольных) шаров по контрасту начинает казаться больше правого, — то оказывается, что иллюзия (называемая «фиксированной установкой») удерживается достаточно длительное время даже при воздействии посторонних интерферирующих раздражителей. То же имеет место и у больных со стертыми нарушениями памяти, возникающими, например, при опухолях гипофиза (Н. К. Киященко, 1969, 1973).
Если этот же опыт провести с больными, у которых в результате массивных поражений глубоких отделов мозга возникают грубые первичные нарушения памяти, результаты оказываются иными. Хорошо удерживая фиксированную установку в течение незаполненного побочными раздражителями интервала времени, они сразу же теряют ее, если этот интервал заполняется побочной деятельностью (Н. К. Киященко, 1969, 1973). Этот факт показывает, что патологическая тормозимость следов распространяется в этих случаях не только на произвольную мнестическую деятельность, но и на все следы, которые не были объектом специального, преднамеренного, запоминания.
Сходные факты можно обнаружить с помощью известного опыта Ю. М. Конорского, заключающегося в том, что испытуемому предъявляют определенный раздражитель (например, окрашенную геометрическую фигуру), который через 30 — 60 — 120 с сменяется вторым — либо идентичным, либо отличающимся (например, геометрическая фигура другой формы или другого цвета). Нормальный испытуемый (как и больной со стертыми поражениями памяти) легко сохраняет образ первой фигуры и четко оценивает тождественность второй фигуры как в условиях «пустого» интервала, так и в условиях интервала, «заполненного» посторонней деятельностью.
У больных с массивными, глубокими поражениями мозга и грубыми нарушениями памяти этот опыт удается лишь тогда, когда интервалы между первым и вторым раздражителями не заполняются побочной деятельностью (Н. К. Киященко, 1969 1973).
Отличие стертых, неспецифических нарушений памяти от грубых форм, наступающих при массивных поражениях глубоких отделов мозга, обнаруживается не только в приведенных выше условиях.
288
Существенное различие только что описанных степеней первичного модально-неспецифического нарушения памяти заключается в том, что если при стертых нарушениях памяти переход к удержанию организованных (осмысленных) мнестических групп оказывается достаточным для компенсации дефекта, то при массивных поражениях памяти того же типа подобный переход к организованным (осмысленным) мнестическим группам не избавляет больного от тормозящего влияния интерферирующих агентов, и нарушения памяти легко превращаются в нарушения сознания.
Все это показывает, что массивные поражения глубин мозга препятствуют адекватному кодированию запоминаемого материала и приводят к таким нарушениям первичной памяти, которые делают недействительным закон Бюлера, согласно которому запоминание мыслей несравненно более прочно, чем запоминание изолированных элементов, указывает на тесную связь этих образований с наиболее грубыми мнестическими расстройствами и стирает грань между нарушениями памяти и расстройствами сознания, которые эти поражения вызывают.
Далее мы остановимся на особых, грубейших формах нарушения смысловой памяти, когда к поражению глубоких подкорковых отделов присоединяется поражение лобных долей мозга.
3 МОДАЛЬНО-СПЕЦИФИЧЕСКИЕ ФОРМЫ ПАМЯТИ
В отличие от неспецифических расстройств памяти, возникающих при поражении аппаратов первого функционального блока мозга, нарушения памяти, возникающие при поражениях конвекситальных отделов коры, иначе говоря, при поражениях второго и третьего функциональных блоков, носят принципиально иной характер. Основная черта этих нарушений заключается в том, что они никогда не являются глобальными и никогда не связаны с общим «расстройством» сознания. Как правило, они либо носят характер нарушения мнестической основы отдельных модально-специфических операций (той или иной стороны познавательных процессов), либо же оказываются связанными с расстройствами динамики активной, целенаправленной деятельности.
Рассмотрим эти виды нарушений.
Типичными для поражения наружных отделов височной доли левого (доминантного у правши) полушария являются упомянутые выше специфические нарушения слухоречевой памяти. Как показало значительное число исследований (А. Р. Лурия, 1940, 1947, 1970; Э.С.Бейн, 1947; М. Климковский, 1966), ни в одном
289
из наблюдавшихся случаев локальное поражение коры левой височной области не приводило ни к общим нарушениям памяти или сознания, о которых мы говорили, описывая больных с глубокими поражениями мозга, ни к той инактивности мнестических процессов, о которых речь пойдет далее при анализе расстройств мнестической деятельности при поражении лобных долей мозга. Лишь в тех случаях, когда поражение распространялось на глубокие (медиальные) отделы височной доли и протекало на фоне выраженных общемозговых изменений, у больного наблюдались также признаки дезориентированности и спутанности сознания, типичные для только что описанных форм поражения глубоких отделов мозга, расположенных по средней линии.
Поражение конвекситальных отделов коры левой височной области приводит прежде всего к распаду сложных речевых форм слухового гнозиса, выражающемуся в дефектах фонематического слуха, и именно следствием этих поражений являются те высокоспецифические нарушения слухоречевой памяти, на которых мы уже останавливались.
В случаях массивных поражений верхневисочных отделов левого полушария эти расстройства маскируются или перекрываются явлениями сенсорной афазии; в случае поражения средних отделов височной области они выступают особенно резко и проявляются в невозможности удержать сколько-нибудь длинные серии звуков или слов, что составляет основной признак акустико-мнестической афазии (М. Климковский, 1966; Л. С. Цветкова, 1972).
Характерно, что этот симптом выступает особенно резко в удержании серий речевых звуков и слов; однако он может проявляться и в удержании комплексных тонов и ритмических структур (М. Климковский, 1966), хотя совсем не отмечается при удержании серий зрительных образов, движений (А.Р.Лурия, М.Ю.Рапопорт, 1962; А.Р.Лурия и др., 1970; Фам Мин Хак, 1971).
Можно предположить, что в основе этих дефектов лежит повышенная тормозимость слухоречевых следов, в результате которой одни элементы слухоречевого ряда либо тормозятся другими, либо всплывают с равной вероятностью с побочными следами (А.Р.Лурия, Е.Н.Соколов, М.Климковский, 1967). Характерно, наконец, что такие дефекты нарушения слухоречевой памяти успешно преодолеваются, если элементы слухоречевого ряда даются больному с относительно большими интервалами (Л. С. Цветкова, 1969).
Таким образом, все описанные дефекты памяти, возникающие при поражениях левой височной области, тесно связаны со слуховыми процессами и речью, иными словами, носят строго модально-специфический характер.
Иными чертами отличаются нарушения памяти, возникающие при поражении левой теменной (или теменно-затылочной) области.
290
Известно, что в этих случаях, как это было уже описано выше, у больного возникают выраженные затруднения симультанных синтезов, и нарушение его мнестических процессов является прямым следствием этих гностических расстройств.
Как показали исследования Фам Мин Хака (1971), запоминание простых фигур (как и звуков) остается в этих случаях незатронутым, но как восприятие, так и удержание зрительных структур, включающих симультанные (пространственные) отношения, протекает с большим трудом и длительная тренировка в их непосредственном запечатлении обычно не приводит к нужным результатам.
Мы уже упоминали, что симптом нарушения припоминания названий предметов, широко известный в клинике под названием амнестической афазии, возникает при поражениях именно этой области коры и, по всей вероятности, является следствием патологически измененной нейродинамики этих корковых отделов, приводящей к равновероятному всплыванию различных систем словесных следов (близких по фонетике, морфологии, лексике), что проявляется с полной отчетливостью в литеральных и вербальных парафазиях (Лотмар, 1919, 1935; И.Д.Сапир, 1929; Э.С.Бейн, 1957; А.Р.Лурия, 1970).
В этих случаях также модально-специфические нарушения памяти остаются дефектами операций запоминания и припоминания, но никогда не превращаются в нарушения структуры самой мнестической деятельности, сохраняя тем самым возможность компенсации этих дефектов (А.Р.Лурия, 1948, 1963; Э.С.Бейн, 1947, 1957; Л.С.Цветкова, 1972).