
1 постцентральные зоны коры и афферентная организация движений








Постцентральные (или кожнокинестетические) отделы коры расположены сзади от центральной борозды и имеют типичное для аппаратов второго блока мозга мелкоклеточное, зернистое строение. В первичных отделах этой области (3-е поле Бродмана), как и во всех проекционных полях, преобладающее место занимает IV афферентный слой коры. Для этих отделов характерно отчетливое соматотопическое строение, причем, как это было указано ранее, волокна, несущие импульсы от нижних противоположных конечностей, располагаются в верхних отделах этой зоны, заходя частично на ее медиальную поверхность; волокна, несущие импульсы от верхних конечностей, — в средних, а волокна, несущие импульсы от лица, губ, языка, — в нижних отделах. Как уже было сказано, эта проекция построена не столько по геометрическому, сколько по функциональному принципу: чем большее значение имеет та или иная область периферических рецепторов и чем большей управляемостью характеризуется соответствующий ей двигательный сегмент, тем большую территорию занимает его проекция в упомянутых зонах коры.
Совершенно естественно, что результатом локального поражения описываемых областей мозга является выпадение (или снижение) чувствительности в соответствующих сегментах тела. Одна-
ко это является лишь наиболее прямым и очевидным, но далеко не единственным результатом такого поражения. Как известно, нормальная кожнокинестетическая афферентация является необходимой основой движения. Она придает двигательным импульсам направленность, нарушаясь при поражении задних, постцентральных, отделов коры и ее проводящих путей, приводя к явлениям своеобразного афферентного пареза, при котором потенциальная сила мышц остается сохранной, но возможность управлять движениями конечности резко снижается и больной оказывается не в состоянии производить тонкие произвольные движения (Ферстер, 1936).
При афферентном парезе двигательные импульсы теряют четкий, дифференцированный адрес и перестают доходить до нужных мышечных групп. Это становится очевидным при сравнении электромиограммы движений сгибания и разгибания у нормального человека (рис. 73, а) и у человека с поражением (опухолью) постцентральной области мозга (рис. 73, б, в). В последнем случае импульсы одновременно «затекают» как в группу агонистов, так и в группу антагонистов, и нужное движение не выполняется.
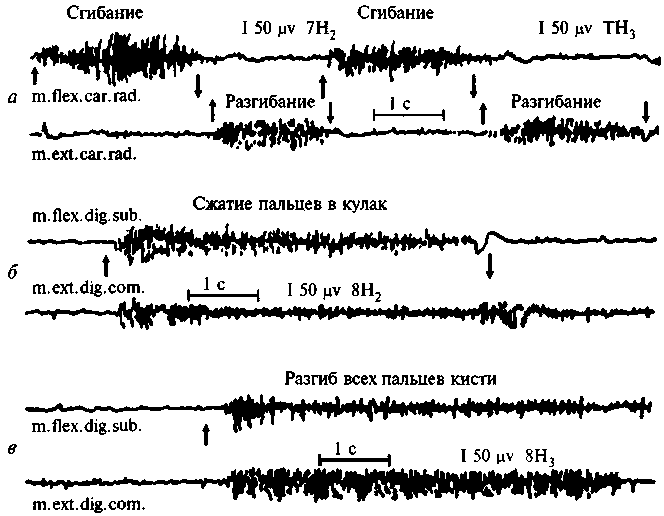
Рис. 73. Электромиограмма движений сгибания и разгибания
(по И.И.Замбран):
а — в норме; б, в — у больного с опухолью постцентральных областей коры
Над первичной, проекционной кожнокинестетической корой, так же как и в случае других модально-специфических отделов этого блока мозга, надстроены вторичные отделы. К ним относятся 1-, 2-, 5-, 7- и частично 40-е поля Бродмана, в которых, как и в других вторичных полях, существенное место занимают верхние слои; эти поля, сохраняя модально-специфический характер, теряют, однако, соматотопическую организацию. Входящие в их состав нейроны отвечают на более комплексные раздражители, а раздражение вторичных кинестетических зон коры вызывает более распространенные ощущения по сравнению с раздражением первичных зон.
При поражении аналогичных отделов постцентральной области грубые нарушения чувствительности обычно отсутствуют, и на первый план выступают нарушения комплексных форм кожнокинестетической чувствительности, проявляющиеся в невозможности синтеза отдельных ощущений в целые структуры. Как следствие у больных нарушаются наиболее сложные формы активного осязания (Б.Г.Ананьев и др., 1959) и отмечаются явления, аналогичные симультанной и предметной оптическим агнозиям, наблюдающимся при патологии вторичных отделов затылочной коры. Эти явления получили в последнее время название дефекта аморфосинтеза (Денни-Браун, 1952, 1958). Этот дефект, по-видимому, лежит в основе хорошо известного в клинике явления астереогноза (невозможности узнавать на ощупь предъявляемые предметы), проявляющегося, как правило, в контралатеральной очагу руке.
177
Было бы неверным думать, что нарушения, возникающие при поражении вторичных отделов постцентральных отделов коры, ограничиваются лишь афферентными или гностическими расстройствами.
Существенной особенностью этих отделов коры является тот факт, что их поражение неизбежно сказывается на протекании двигательных процессов.
Как уже отмечалось, организованное протекание произвольного (предметного) движения в высокой степени зависит от системных кинестетических афферентаций (Н. А. Бернштейн, 1947; и др.). Поэтому естественно, что если кинестетические синтезы нарушаются, исчезает и непосредственная афферентная основа движения. Вот почему при поражении вторичных отделов постцентральной кинестетической коры у больного возникают своеобразные явления афферентной апраксии, которые заключаются в том, что рука, не получающая нужных афферентных (кинестетических) сигналов, не может выполнить тонкие дифференцированные движения. Явления афферентной апраксии были в свое время описаны Липманом (1905, 1920), который обозначил их как «акро-кинетическую апраксию», затем Клейстом (1907, 1911), А. Пиком (1905), Монаковым (1905, 1914), Брунсом (1921), Зиттигом (1931). Наиболее полное описание их принадлежит Ферстеру (1936), показавшему, что рука больного с поражением вторичных отделов постцентральной области мозга лишается возможности адекватно приспосабливаться к характеру предмета и превращается, по выражению Ферстера, в «руку-лопату» (рис. 74). Такие нарушения, называемые также «апраксией позы», или «афферентной или кинестетической апраксией» (А. Р.Лурия, 1963, 1969), можно считать достаточно надежным признаком поражения вторичных отделов кинестетической, постцентральной коры.
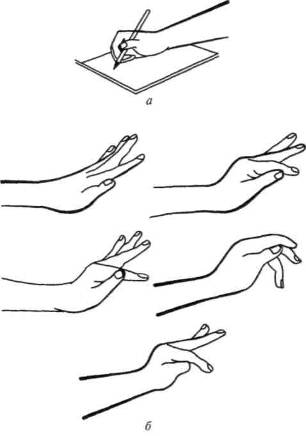
Рис. 74. Нарушение движений руки при поражении
постцентральной области (по Форстеру):
а — нарушение тонких движений при письме; б — явление статической атаксии
Необходимо, однако, специально рассмотреть одно обстоятельство. Если поражение вторичных (кинестетических) отделов постцентральной коры левого (доминантного) полушария распространяется на нижние отделы этой области (области вторичной организации кинестезии лица, губ и языка), кинестетическая апрак-сия может проявиться в нарушении организации движений речевого аппарата, влекущем за собой своеобразные расстройства речи, известные под названием афферентной моторной афазии (А. Р.Лурия, 1947; Е. Н.Винарская, 1969, 1970). Основным в этом синдроме является невозможность найти положения губ и языка, необходимые для артикуляции соответствующих звуков речи; больные с массивной афферентной моторной афазией не знают, какое положение следует придать языку и губам для произнесения нужных звуков; больные с более стертой формой такого нарушения смешивают близкие (отличающиеся каким-нибудь одним признаком) артикулемы; они заменяют нёбно-язычные артикуляции, произнося «д» вместо «л», или путают близкие губные артикуляции, заменяя «б» или «п» на «м». Смешение звуков, разных по акустическим качествам, но близких по артикуляции, может служить верным опорным признаком для диагностики поражений нижних постцентральных отделов левого полушария.
Вторичное (системное) влияние этого дефекта проявляется в своеобразных нарушениях письма, также состоящих в смешении близких артикулем (типа «л»—«н»—«д») — больные могут написать «ха-дат» или «ханат» вместо «халат», «енот» или «слон» вместо «стол» и т.д. (рис. 75); такая замена близких по артикуляции звуков отличает их от больных с поражением левой височной области, в письме которых на первый план выступает смешение близких фонем.

179
Все описанные расстройства являются результатом нарушения афферентной основы движения. Они характеризуют одну сторону патологии движений при локальных поражениях мозга и отличаются от расстройств, сопровождающих поражения премоторных отделов мозга.
2 ПРЕМОТОРНЫЕ ЗОНЫ КОРЫ И ЭФФЕРЕНТНАЯ ОРГАНИЗАЦИЯ ДВИЖЕНИЙ
Постцентральные зоны мозговой коры оказывают настраивающее, модулирующее влияние на отделы мозга, известные как первичные, проекционные двигательные зоны коры больших полушарий. Эти зоны, расположенные в пределах передней центральной извилины и располагающие аппаратом мощно развитых ги-
180
гантских пирамидных клеток V афферентного слоя, являются началом двигательного пирамидного пути, по которому импульсы направляются к передним рогам спинного мозга, а затем — к соответствующим мышечным группам. Они имеют четкое соматотопическое строение подобно общечувствительным (кожнокине-стетическим) зонам задней центральной извилины и находятся под постоянным влиянием этих последних, приводя структуры возникающих в них двигательных импульсов в соответствие с динамическими структурами возбуждений, возникающих в постцентральных отделах мозга.
Афферентные, постцентральные отделы мозга не являются, однако, единственными аппаратами, которые оказывают на переднюю центральную извилину управляющее, модулирующее влияние.
Движение является процессом, протекающим во времени и предполагающим наличие цепи сменяющих друг друга импульсов. На начальных ступенях формирования движения эта цепь состоит из серии изолированных импульсов; по мере формирования двигательных навыков отдельные импульсы синтезируются, объединяются в целые кинетические структуры, или кинетические мелодии, и тогда одного импульса достаточно, чтобы вызвать целый динамический стереотип автоматически сменяющих друг друга звеньев. В выработке такого динамического стереотипа и состоит формирование двигательного навыка, который в результате упражнений становится таким же автоматизмом, как и элементарные врожденные автоматизмы, осуществляющиеся на подкорковом уровне, названном Н.А. Бернштейном (1947) уровнем «двигательных синергии».
У низших позвоночных уровень подкорковых синергии включает в свой состав аппараты зрительного бугра и подкорковых двигательных узлов («таламостриальную систему»); у высших позвоночных над этим уровнем надстраиваются аппараты мозговой коры, наиболее важное место среди которых занимают аппараты премоторных отделов коры головного мозга (см. схему связей этих отделов на рис. 76). По своему строению премоторная область коры (6-е и 8-е поля Бродмана) близка к двигательной коре (вертикальная исчерченность), но в отличие от первичной двигательной зоны она характеризуется большим развитием мелких пирамид II и III слоев, делающим ее типичной вторичной кортикальной зоной.
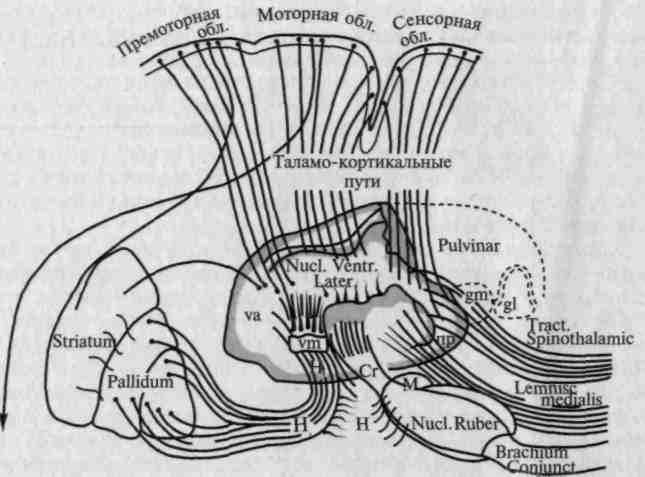
Рис. 76. Схема связей подкоркового и коркового уровней, осуществляющих чувствительные и двигательные поиски
Отметим, что премоторная зона мозговой коры наиболее мощно развивается на последних этапах эволюции млекопитающих; если у низших обезьян преобладает первичное (4-е) двигательное поле, а премоторные отделы коры занимают незначительное место, то у человека это отношение меняется на обратное и премоторная зона коры занимает подавляющую часть прецентральной области (табл. 12).
Таблица 12
Относительная площадь, занимаемая прецентральной (двигательной)
и премоторной областями на последовательных ступенях эволюции
высших млекопитающих
(по данным Московского института мозга, 1958)
| Вид | Отношение | ||
| площади прецентральной области к общей поверхности коры | площади первичной двигательной коры (поля 4) к площади всей прецентральной области | площади премоторной области (поля 6) к площади всей прецентральной области | |
| Игрунка | 5,5 | 79 | 21 |
| Мартышка | 8,3 | 69 | 31 |
| Шимпанзе | 7,6 | 29,8 | 70,2 |
| Человек | 8,4 | 12,0 | 88 |
Таблица 13
Общее число клеток в отдельных слоях первичных, вторичных и третичных полей двигательной коры человека, млн
{по данным Московского института мозга, 1960)
| Поле мозговой коры | Общее количество | Слои коры | ||
| Н-Ш | IV | V | ||
| 4 | 520 | 30 | — | 10,6 |
| 6 | 511 | 275 | — | 80,6 |
| 10 | 917 | 386 | 94 | 177 |
181
Анатомо-физиологический и критический анализы позволяют существенно уточнить характер участия премоторных зон в двигательных процессах. Как показали морфологические исследования, у человека в первичной двигательной зоне (4-е поле Бродмана) клетки в основном сконцентрированы в V (эфферентном) слое коры, в то время как количество клеток во II и III («ассоциативных») слоях коры сравнительно невелико, во вторичных, премоторных отделах двигательной коры (6-е поле Бродмана) число клеточных элементов в верхних слоях коры резко возрастает. В наиболее сложных — третичных областях коры (10-е поле Бродмана) — число клеточных элементов еще больше увеличивается (табл. 13).
Премоторная зона не имеет соматотопического строения, которым характеризуется проекционная зона двигательной коры, и ее раздражение, распространяясь на относительно большие территории (рис. 26), вызывает не изолированные сокращения мышечных групп, а комплексные движения, включающие повороты глаз и головы, а затем и туловища в противоположную сторону. Такие же адверзивные эпилептические припадки наблюдаются при наличии постоянных раздражающих факторов (например, рубцов) в премоторной области.
182
Итак, премоторные отделы коры являются аппаратом, приспособленным для интеграции эфферентных {двигательных) импульсов, и если постцентральные отделы коры обеспечивают пространственное распределение двигательных импульсов, то премоторные зоны коры, опираясь на нижележащие аппараты двигательных синергии, осуществляют превращение отдельных двигательных импульсов в последовательные кинетические мелодии, обеспечивая тем самым второй необходимый аспект организации сложных движений и двигательных навыков.
183
Сказанным выше определяются симптомы, возникающие при поражении премоторной зоны. Как это было показано еще в классических работах Фултона (1935, 1943), а затем и ряда других неврологов (Ферстер, 1936; Клейст, 1934; А.Р.Лурия, 1943, 1947, 1962, 1963, 1969), поражения премоторных отделов мозга не вызывают ни параличей, ни парезов противоположных конечностей. Их основным симптомом является отчетливое нарушение двигательных навыков {skilled movements). Клинически это проявляется в том, что у человека меняется почерк, каждый штрих буквы начинает требовать специального усилия, машинистка теряет быстроту и плавность своей работы, музыкант — плавность выполнения мелодий, квалифицированный рабочий оказывается не в состоянии автоматически выполнять серию операций, входящих в привычный двигательный акт.
Физиологически этот дефект выступает в том, что плавный процесс смены иннервации и денерваций нарушается; электромиографические импульсы, ведущие к возникновению движения, становятся патологически затянутыми (рис. 77).
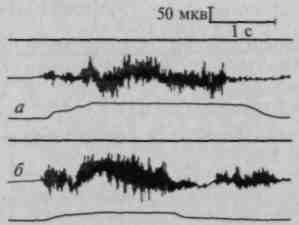
Рис. 77. Патологические изменения электромио- и механограммы при поражении премоторной области:
а — патология начала движения (трудности включения); б — патология конца движения (патологическая инертность) (по А.Я.Иошпа и Е.Д.Хомской)
В клинике такие явления хорошо известны под названием инертности двигательных стереотипов (рис. 78). Если предложить больному с поражением премоторной области отстучать сложный ритм, например ритм, включающий два сильных и три слабых удара, больной оказывается не в состоянии плавно переключаться с сильных ударов на слабые, его отстукивание приобретает отрывистый, дезавтоматизированный характер (А. Р. Лурия, 1943, 1963; Ф. М. Семерницкая, 1945; Б.Г.Спирин, 1966; и др.).
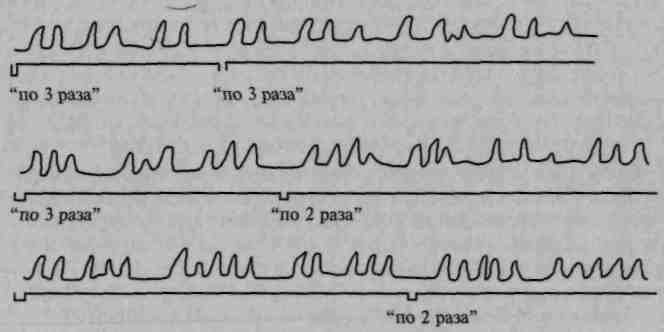
Рис. 78. Инертность элементарных двигательных стереотипов при
выполнении ритмических движений больным с поражением
премоторной области
В тех случаях, когда поражение премоторной области располагается глубоко, наблюдается иной симптом, имеющий большое принципиальное и диагностическое значение: тормозящая и модулирующая функции премоторной коры в отношении нижележащих подкорковых образований (подкорковых двигательных уз-
лов) устраняются, и начавшийся элемент движения не тормозится вовремя, бесконтрольно продолжаясь. Возникает явление, которое Н.А. Бернштейн (1947) назвал «зацикливанием» движения и которое широко известно под названием элементарной двигательной персеверации. На рисунках 79 и 80 приведены примеры таких нарушений движения.

Рис. 79. Двигательные персеверации у больного после пункции премоторной области, сопровождавшейся кровоизлиянием: а — рисунки и цифры; б — письмо

Рис. 80. Двигательные персеверации в послеоперационный период у больного с удалением опухоли (менигеомы) премоторной области:
а — рисование фигур (2—5-й дни после операции); б — рисование человечка (те же дни); в — рисование фигур и человечка на 7-й день после операции
Следует отметить, что наиболее отчетливо описываемые нарушения проявляются в контралатеральной очагу руке, но при поражении премоторной зоны левого (доминантного) полушария они часто проявляются в работе обеих рук. Отметим также, что во всех случаях у больных сохранны как намерение выполнить движение, так и общий план его выполнения, однако в своей исполнительной части движение высвобождается из-под влияния программы.
Остановимся еще на одном важном симптоме. В тех случаях, когда поражение располагается в нижних отделах премоторной зоны левого (доминантного) полушария, только что описанные нарушения плавного переключения с одного двигательного звена на другое (двигательные персеверации) начинают проявляться не только в движениях руки, но и в речи больного, приводя к своеобразному явлению, которое мы обозначили термином эфферентная (или кинетическая) моторная афазия (А. Р.Лурия, 1947, 1962, 1969, 1970).
В отличие от описанной выше афферентной (кинестетической) афазии нахождение нужных артикуляций и произношение изолированных речевых звуков не представляет для этих больных сколько-нибудь заметных трудностей. Отчетливые нарушения наступают тогда, когда больные переходят от одной артикуляции к другой (что имеет место при произношении любого слова или словосочетания). Процесс денервации предыдущей артикулемы и плавного переключения на последующую оказывается глубоко нарушенным, возникают явления патологической инертности раз возникшей артикуляции. Пытаясь произнести слово «муха», такой больной правильно артикулирует (губной) согласный, входящий в первый слог «му», но оказывается не в состоянии перейти на следующий (заднеязычно-гортанный) согласный в слоге «ха», и вместо требуемого слова произносит: «му...-м...м...му...ма».
Это нарушение кинетической организации артикуляций является основой той формы моторной афазии, которую в свое время описал Брока (1861) и которая по своим физиологическим механизмам резко отличается от описанной выше апрактической афферентной моторной афазии.
Следует отметить, что подобные нарушения выступают у больных с поражением нижних отделов левой премоторной зоны не только в устной речи, но и в письме как следствие нарушения плавности перехода от одного компонента слова к другому и патологической персеверации раз написанного слова (рис. 81).
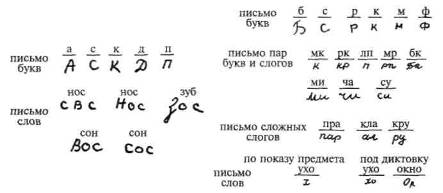
Рис. 81. Письмо больных с эфферентной (кинетической) моторной афазией
187
Таким образом, изучение функций премоторных отделов коры дает нам возможность вплотную подойти к механизмам одного из наиболее сложных явлений — нарушений моторной стороны речи.
Глава V
ЛОБНЫЕ ДОЛИ МОЗГА И РЕГУЛЯЦИЯ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ
Премоторные отделы мозговой коры входят в третий, основной, блок мозга, обеспечивающий программирование, регуляцию и контроль человеческой деятельности.
Как известно, лобные доли мозга, и в частности их третичные образования (к которым относится префронтальная кора), являются наиболее поздно сформировавшимся отделом больших полушарий. Едва намеченная у высших животных префронтальная кора заметно увеличивается у приматов, а у человека занимает до 25 % всей площади больших полушарий.
Префронтальные отделы мозга — или лобная гранулярная кора — состоят, главным образом, из клеток верхних (ассоциативных) слоев коры. Они имеют богатейшие связи как с верхними отделами ствола и образованиями зрительного бугра (рис. 45, а), так и со всеми остальными зонами коры (рис. 45, б). Таким образом, префронтальная кора надстраивается не только над вторичными отделами двигательной области, но фактически над всеми остальными образованиями большого мозга. Это обеспечивает двустороннюю связь префронтальной коры как с нижележащими структурами ретикулярной формации, модулирующими тонус коры, так и с теми образованиями второго блока мозга, которые обеспечивают получение, переработку и хранение экстероцептивной информации, что позволяет лобным долям регулировать общее состояние мозговой коры и протекание основных форм психической деятельности человека. Обе эти функции лобных долей мозга имеют настолько важное значение, что мы рассмотрим каждую из них по отдельности.
Функция лобных долей в организации поведения была подробно изучена многими авторами. Анализу их роли в организации поведения животных посвящены классические работы В.М.Бехтерева (1907), И.П.Павлова (1912-1913), П.К.Анохина (1949), Бианки (1895, 1921), Франца (1907), Джекобсона (1935), Малмо (1942), Прибрама (1954-1960), Росволда (1956 — 1959), Мишкина (1955—1958) и др. Анализ роли лобных долей в поведении человека был осуществлен в исследованиях Харлоу (1868), Вельт (1888), В.К.Хорошко (1912, 1921), Фейхтвангера (1923), Клейста (1943), Брикнера (1936), Райлендера (1939), Хэбба (1945), Халстеда (1947), Денни-Брауна (1951), А.Р.Лурия (1962, 1963, 1969), А.Р.Лурия, Е.Д.Хомской (1964, 1966), Е.Д.Хомской (1972) и др.
В следующей главе будут коротко изложены те данные, которые были получены в этих исследованиях.
189