
Рис. 19. Последовательное увеличение размеров первичных, вторичных и третичных зон коры в онтогенезе (по данным Московского института мозга)









Лурия А. Р.
Основы нейропсихологии. Учеб. пособие для студ. высш. учеб. заведений. — М.: Издательский центр «Академия», 2003. — 384 с.
УДК 57.024(075.8)
ББК 88.4я73
Л 86
ISBN 5-7695-1013-7
Серия «Классическая учебная книга»
Автор предисловия Е.Д. Хомская
Эта книга, издаваемая к 100-летию со дня рождения А. Р. Лурия, предназначена для студентов в качестве учебного пособия еще самим автором. Она построена как учебный курс, в котором кратко излагается содержание нейропсихологии как учебной дисциплины. Автор подробно анализирует психологическую структуру и мозговые механизмы отдельных психических процессов - восприятия, произвольных движений и действий, внимания, памяти, речи, мышления. Раскрывает популярную сегодня среди нейропсихологов и клиницистов модель трех основных блоков мозга, в которой реализованы представления автора о целостном характере деятельности мозга при осуществлении психических процессов и об основных типах их нарушений.
Может быть интересна нейропсихологам разных поколений, а также медикам - невропатологам, нейрохирургам, психиатрам.
© Правопреемник Радковская Е.Г., 2002
© Предисловие. Хомская Е.Д., 2002
© Издательский центр «Академия», 2002
ОГЛАВЛЕНИЕ
Предисловие к второму изданию. Е.Д.Хомская.........................................3
От автора.......................................................................................................6
Часть первая
ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ МОЗГА И ПСИХИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ (основные принципы)
Введение.......................................................................................................8
Глава I. Три источника знаний о функциональной организации мозга.......11
1. Сравнительно-анатомические данные.......................................11
Основные принципы эволюции и строения мозга как органа психики...............................12
Структурная и функциональная организация коры головного мозга..................................22
2. Физиологические данные: метод раздражения.........................45
Опыты с непосредственным раздражением коры....................45
Опыты с непрямой стимуляцией коры.....................................53
Опыты с анализом функций отдельных нейронов...................57
3. Физиологические данные: метод разрушения...........................60
Глава II. Локальные поражения мозга и основные принципы локализации функций...................................................................67
1. Ранние решения...........................................................................67
2. Кризис..........................................................................................72
3. Пересмотр основных понятий....................................................74
Пересмотр понятия «функция»..................................................74
Пересмотр понятия «локализация»............................................77
Пересмотр понятия «симптом»...................................................80
4. Синдромный анализ и системная организация психических процессов......................................................................................84
Глава III. Три основных функциональных блока мозга............................88
1. Блок регуляции тонуса и бодрствования...................................88
2. Блок приема, переработки и хранения информации.............101
3. Блок программирования, регуляции и контроля сложных форм деятельности.....................................................................111
4. Взаимодействия трех основных функциональных блоков мозга...........................126
Часть вторая
ЛОКАЛЬНЫЕ СИСТЕМЫ МОЗГА И ИХ ФУНКЦИОНАЛЬНЫЙ АНАЛИЗ
Вступительные замечания........................................................................130
Глава I. Затылочные отделы мозга и организация зрительного восприятия..................................................................................132
1. Первичные зоны затылочной коры и элементарные функции зрения.........................................................................132
2. Вторичные отделы затылочной коры и оптико-гностические функции......................................................................................135
Глава II. Височные отделы мозга и организация слухового восприятия..................................................................................149
1. Первичные зоны височной коры и элементарные функции слуха...................149
2. Вторичные отделы височной коры и акустико-гностические функции......................................................................................151
3. Системное влияние нарушений речевого слуха на другие психические процессы..............................................................156
4. Варианты «височного синдрома».............................................159
Глава III. Третичные зоны коры и организация симультанных синтезов.......................................................................................163
1. Третичные зоны коры и организация наглядных пространственных синтезов.....................163
2. Третичные зоны коры и организация символических (квазипространственных) синтезов.........................................167
3. Третичные зоны коры и процессы речевой памяти...............170
4. Теменно-затылочные зоны правого (субдоминантного) полушария и их функции..........................................................173
Глава IV. Сенсомоторные и премоторные отделы мозга и организация движений............................................................175
1. Постцентральные зоны коры и афферентная организация движений....................................................................................175
2. Премоторные зоны коры и эфферентная организация движений....................................................................................180
Глава V. Лобные доли мозга и регуляция психической деятельности......189
1. Лобные доли и регуляция состояний активности...................190
2. Лобные доли и регуляция движений и действий....................199
3. Лобные доли и регуляция мнестических и интеллектуальных действий.............................209
4. Функциональная организация лобных долей и варианты лобного синдрома......................................................................216
Глава VI. Медиобазальные отделы коры, правое полушарие мозга........220
1. Медиальные отделы коры и их роль в регуляции психических состояний.....................................220
2. Субдоминантное полушарие и его роль в организации психических процессов..............................................................225
Часть третья
ПСИХИЧЕСКИЕ ПРОЦЕССЫ И ИХ МОЗГОВАЯ ОРГАНИЗАЦИЯ
Глава I. Восприятие..................................................................................232
1. Психологическое строение........................................................232
2. Мозговая организация...............................................................233
Глава П. Движение и действие.................................................................246
1. Психологическое строение........................................................246
2. Мозговая организация...............................................................251
Глава III. Внимание...................................................................................256
1. Психологическое строение........................................................256
2. Физиологические индикаторы внимания................................263
3. Мозговая организация...............................................................268
Глава IV. Память.......................................................................................276
1. Психологическое строение........................................................276
2. Модально-неспецифические формы памяти..........................282
3. Модально-специфические формы памяти..............................289
4. Нарушение памяти как мнестической деятельности..............291
Глава V. Речь.............................................................................................294
1. К истории вопроса.....................................................................294
2. Психологическое строение речевой деятельности..................296
3. Импрессивная речь....................................................................298
4. Экспрессивная речь...................................................................301
Глава VI. Мышление.................................................................................309
1. Психологическое строение........................................................309
2. Наглядное (конструктивное) мышление.................................315
3. Вербально-логическое (дискурсивное) мышление. Решение задач............................................................................................319
Заключение................................................................................................324
Литература.................................................................................................328
Указатель имен..........................................................................................364
Предметный указатель..............................................................................368
ПРЕДИСЛОВИЕ
к второму изданию
Первое издание этой книги на русском языке вышло почти 30 лет тому назад (в 1973 г.). Это единственная книга А. Р.Лурия, специально предназначенная для студентов в качестве учебного пособия. Все остальные книги — научные монографии или сборники работ под его редакцией. Однако деление книг А. Р.Лурия на учебные и научные довольно условно. «Основы нейропсихологии» — одна из важнейших классических работ А. Р.Лурия, имеющая прежде всего большое научное значение. Вместе с тем многие его научные монографии используются и в учебных целях для подготовки как нейропсихологов, так и клинических психологов широкого профиля.
Первоначально А. Р.Лурия хотел назвать эту книгу «Работающий мозг» (или «Активный мозг»), но затем изменил свое намерение. Однако за рубежом она вышла под названием « The working brain: an introduction to neuropsychology» (New York, 1973). Выбор названия, конечно, неслучаен. В нем подчеркивается основная идея книги — показать, как именно работает мозг человека во время осуществления той или иной психической деятельности.
«Основы нейропсихологии» — одна из поздних книг А. Р.Лурия. Она была написана позже, чем «Высшие корковые функции» (1962, 1969), «Мозг и психические процессы» (1963. — Т. 1; 1970. — Т. 2), «Маленькая книжка о большой памяти» (1968), «Потерянный и возвращенный мир» (1971). Поэтому в ней суммируются различные сведения по нейропсихологии, как уже устоявшиеся, так и новые, полученные за последние годы. В связи с этим представляется важным сравнить «Основы нейропсихологии» с другими, более ранними книгами А. Р.Лурия, и прежде всего «Высшими корковыми функциями». Различия очевидны.
Книга построена как учебный курс, в котором в относительно краткой форме излагается содержание нейропсихологии как новой научной дисциплины. В ней представлены «обе ипостаси» нейропсихологии, а именно: ее принадлежность к психологическим дисциплинам («психика и мозг») и к нейронаукам («мозг и психика»). В других работах преобладает какой-либо один аспект нейропсихологии. Так, в «Высших корковых функциях» описываются главным образом целостные нейропсихологические синдромы, возникающие при поражении различных областей мозга; показывается комплексный характер нарушений психических функций при локальных очагах. Иными словами, логика изложения материала состоит прежде всего в описании роли тех или иных областей мозга в реализации психических процессов («мозг и психика»).
3
В «Основах нейропсихологии» А. Р. Лурия впервые в систематическом виде представил другой аспект нейропсихологии. Он подробно анализирует психологическую структуру и мозговые механизмы отдельных психических процессов (восприятия, произвольных движений и действий, внимания, памяти, речи, мышления). Эта логика изложения материала («психика и мозг») особенно важна для преподавания курса нейропсихологии, так как она конкретизирует на различных психических процессах суть понимания ученым проблемы локализации высших психических функций.
В книгу включен новый (для 1973 г.) материал, ранее не излагавшийся в других работах. Это — глава «Три основных блока мозга», вошедшая в раздел «Функциональная организация мозга и психическая деятельность». Она является первым подробным описанием общей структурно-функциональной модели работы мозга как субстрата психических процессов. Ранее идея блоков мозга встречалась лишь однажды в докладе А. Р. Лурия на XVI Международном конгрессе по прикладной психологии в Амстердаме (1968). Как известно, модель трех блоков теперь очень популярна среди нейропсихологов и клиницистов: в ней реализованы представления А. Р. Лурия о целостном характере деятельности мозга при осуществлении психических процессов, с одной стороны, и об основных типах их нарушений (в соответствии с поражением того или иного блока) — с другой.
Представлены также новые материалы о функциях правого полушария и медиобазальных отделов мозга, полученные в его коллективе лишь в 70-е годы (работы Э. Г. Симерницкой, Л. И. Московичюте и др.). Эти направления исследований получили интенсивное развитие в современной нейропсихологии. Вообще в «Основах нейропсихологии» А. Р. Лурия четко очерчивает границы уже достигнутого и того, что, с его точки зрения, предстоит сделать в ближайшее время. К первой категории сведений он относит все те знания, которые уже накоплены в нейропсихологии и на которых базируется теория системной динамической локализации высших психических функций. К второй — сведения, относящиеся к таким проблемам, как роль глубоких структур мозга в протекании психических процессов, мозговые механизмы эмоциональных явлений, особенности работы субдоминантного (правого для правшей) полушария мозга и ряд других. Именно эти направления нейропсихологии стали интенсивно разрабатываться в послелуриевский период и являются очень актуальными и в настоящее время.
4
В целом, несмотря на относительно большой срок, прошедший со времени издания «Основ нейропсихологии», их содержание ничуть не устарело. Общая концепция, все основные положения этой книги справедливы и актуальны, как и прежде. Можно отметить лишь, что за эти годы в нейропсихологии появились новые направления (нейропсихология детского возраста, нейропсихология нормы, нейропсихология эмоции др.); в то же время получили развитие и старые направления, что говорит о жизнеспособности и продуктивности созданной А.Р.Лурия нейропсихологической школы и большой эвристичности его идеи [1].
1 Подробно см.: Нейропсихологическая школа А. Р.Лурия // Вопросы психологии. — 1997. — № 5.
Переиздание «Основ нейропсихологии» очень своевременно. Несмотря на сравнительно большой тираж первого издания (22 тыс. экз.), в настоящее время эта книга стала раритетом. Потребность в ней особенно возросла за последние годы, когда началась массовая подготовка клинических психологов в различных учебных заведениях России. «Основы нейропсихологии» относятся к числу важнейших учебных пособий по нейропсихологии, а курс нейропсихологии является одним из основных в цикле дисциплин, входящих в специальную клиническую психологию.
Как известно, в нашей стране клинические психологи, в том числе и нейропсихологи, готовятся на базе не только психологического образования, но и других видов специального образования (медицинского, педагогического, даже технического). За последние годы особенно велик приток в клиническую психологию лиц, не имеющих систематических психологических знаний (больше всего их среди тех, кто получает второе образование). Для таких студентов книга «Основы нейропсихологии» особенно нужна, так как в ней наряду с нейропсихологическими знаниями содержатся и общепсихологические. Как известно, А. Р. Лурия строил нейропсихологию на основе общепсихологических концепций и рассматривал ее как «приложение» этих концепций к мозгу. Этот общепсихологический «каркас» повышает ценность «Основ нейропсихологии» как учебного пособия, так как он формирует у студентов профессиональное психологическое мировоззрение, необходимое для любой так называемой прикладной психологии, в том числе и клинической.
«Основы нейропсихологии» публикуется в год, когда исполняется 100 лет со дня рождения А. Р. Лурия (1902 — 1977), и накануне II Международной конференции, посвященной этой дате. Новое издание этой книги будет прекрасным подарком для всех почитателей А.Р.Лурия, и особенно для нейропсихологов разных поколений, включая и будущих специалистов.
Профессор Е.Д. Хомская
5
ОТ АВТОРА
Предлагаемая книга является введением в нейропсихологию — новую отрасль психологии и медицины, сложившуюся за последние тридцать лет при ближайшем участии автора и его сотрудников. Задача нейропсихологии — это изучение мозговых основ психической деятельности человека с привлечением новых, психологических, методов для топической диагностики локальных поражений мозга. Именно это позволяет думать, что данная книга окажется полезной как психологам, так и медикам, и в первую очередь невропатологам, нейрохирургам и психиатрам.
Нейропсихология — молодая наука; естественно, что ее различные разделы разработаны неодинаково. Этим объясняется тот факт, что в книге отсутствует ряд разделов, по которым у автора недостаточно собственного материала и которые он предпочел оставить за пределами обсуждения. К ним относятся разделы, посвященные роли глубоких (в частности, гипоталамических и таламических) структур в протекании психических процессов, мозговым механизмам сна и бодрствования, мозговым основам эмоциональной жизни, а также значению субдоминантного (правого) полушария для психической деятельности человека.
Как и при подготовке предшествующих книг, большую помощь автору оказал коллектив Института нейрохирургии им. Н. Н. Бурденко Академии медицинских наук СССР и сотрудники руководимой автором кафедры нейро- и патопсихологии факультета психологии Московского государственного университета. Всем им — и особенно многолетнему сотруднику доктору психологических наук Е. Д.Хомской, давшей ряд ценных советов и взявшей на себя труд отредактировать рукопись этой книги, — автор приносит искреннюю благодарность.
А.Р.Лурия
6
Часть первая
ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ МОЗГА И ПСИХИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ
(ОСНОВНЫЕ ПРИНЦИПЫ)
ВВЕДЕНИЕ
Мозг как орган психической деятельности в настоящее время стал средоточием научных интересов ряда дисциплин.
Как построен мозг человека, этот совершенный прибор, позволяющий осуществлять сложнейшие формы отражения действительности, и какова его функциональная организация? Какие аппараты мозга обеспечивают возникновение специфических потребностей и намерений, отличающих человека от животного? Как организованы нервные процессы, связанные с получением, переработкой и хранением информации, поступающей из внешнего мира? Чем обеспечиваются программирование, регуляция и контроль наиболее сложных форм сознательной деятельности, направленной на достижение целей, осуществление намерений и реализацию планов?
Еще несколько десятилетий назад эти вопросы не стояли сколько-нибудь остро. Наука вполне удовлетворялась уподоблением мозга реагирующим приборам, ограничиваясь в его познании элементарными схемами, объединявшими стимулы, приходящие из внешнего мира, и обусловленные прошлыми воздействиями ответы на эти стимулы.
К настоящему времени положение коренным образом изменилось. Стало совершенно ясно, что поведение человека носит активный характер, что оно определяется не только прошлыми воздействиями, но и планами, и намерениями; не только создает соответствующие модели будущего, но и подчиняет им поведение. Стало вместе с тем очевидно, что замыслы и намерения человека, схемы будущего и реализующие их программы не должны оставаться вне сферы научного знания и что лежащие в их основе механизмы могут и должны стать предметом такого же детерминистического анализа и научного объяснения, как и все другие явления и связи объективного мира.
Тенденция к изучению механизмов влияния будущего на реальное поведение вызвала к жизни ряд важнейших физиологических концепций, например концепции «опережающего возбуждения» П. К. Анохина и «двигательной задачи и ее реализации» Н.А. Бернштейна, что явилось признаком коренной смены интересов в физиологической науке, основной задачей которой стало теперь создание «физиологии активности».
Радикально изменился и основной теоретический смысл науки о мозге. Если ранее теория мозга основывалась на механических представлениях (моделях) и допускала возможность объяснения работы мозга, исходя из принципов построения телефонной станции или пульта управления, то в настоящее время мозг человека рассматривается как сложнейшая и своеобразно построенная функциональная система, работающая по специфическим принципам, знание которых может помочь исследователям в построении новых математических и реально действующих схем, позволяющих приблизиться к созданию механических аналогов этого совершенного органа.
8
Вот почему изучение внутренних закономерностей работы мозга — как бы трудно ни было их познание — привело к возникновению совершенно новых научных дисциплин. Одна из них — бионика — непосредственно предполагает изучение мозга как источника познания новых принципов, которые оказали бы влияние на творческое развитие математики и техники.
Изучение законов работы мозга как органа психической деятельности — сложнейшая задача. Поэтому совершенно естественно, что она не может быть решена умозрительным конструированием, которое может лишь скомпрометировать эту важную отрасль науки и, создав видимость решения сложнейших проблем, фактически стать препятствием для ее прогресса. Именно поэтому ряд книг, посвященных проблемам моделей мозга или мозгу как вычислительному устройству, не помогает, а, скорее, препятствует продвижению подлинно научных знаний о мозге как органе психики.
Подлинный прогресс в этой важной области должен опираться не на логические схемы, а на реальные факты, реальные достижения, на результаты кропотливых наблюдений, относящихся к разным областям науки: морфологии и физиологии, психологии и неврологии.
Естественно, что прогресс этот требует времени и что проникновение в неизвестное — длительный процесс, каждый отдельный этап которого вносит свой вклад в окончательное решение поставленных задач.
Около четверти века назад появилась известная книга Грея Уолтера «Живой мозг», в которой была сделана попытка привлечь данные электрофизиологии для объяснения интимных механизмов работы человеческого мозга и высказаны гипотезы (частично подтвердившиеся, частично оставшиеся предположениями автора) об основных формах жизни мозга и принципах его функционирования.
Через несколько лет после этого появилась монография выдающегося анатома и физиолога Г. Мэгуна «Бодрствующий мозг», представлявшая собой попытку рассмотрения мозга на основании новейших анатомических и нейрофизиологических данных как системы, способной к самостоятельному обеспечению бодрствующего, активного состояния, являющегося условием всякого поведения живого существа.
Значение книги Мэгуна, обобщающей достижения целой группы блестящих исследователей — Моруцци, Джаспера, Пенфилда и других, нельзя переоценить. С ее появлением мозг человека и животных перестал расцениваться как пассивно реагирующий аппарат и был сделан первый шаг в познании его как саморегулирующейся системы.
Однако, описав механизмы бодрствования, Мэгун не сделал анализа основных форм конкретной психической деятельности человека. Механизмы познавательной деятельности (восприятия и мышления), речи и общения, формирования планов и программ поведения, регуляции и контроля — весь этот круг проблем остался за рамками книги. Факты, которые позволили бы подойти к решению этих вопросов и создать основы учения о мозге как органе конкретной психической деятельности, постепенно накапливались различными областями науки.
9
Подход к анализу этих фактов наметился благодаря успехам современной научной психологии, описавшей строение человеческой деятельности и вплотную подошедшей к анализу функциональной структуры восприятия и памяти, мышления и речи, движения и действия и процессов их формирования в онтогенезе.
Большое число фактов накопилось в современной неврологической и нейрохирургической клинике, где было детально изучено то, как нарушаются сложнейшие формы поведения при локальных поражениях мозга.
Решение этих вопросов существенно приблизилось созданием новой отрасли науки — нейропсихологии, которая впервые сделала целью научного исследования изучение роли отдельных систем головного мозга в осуществлении психической деятельности.
Все это сделало возможным (и необходимым) подготовку настоящей книги, в которой автор попытался обобщить современные представления о мозговых основах сложной психической деятельности человека и рассказать о том, какие системы головного мозга принимают участие в построении восприятия и действия, речи и мышления, движения и целенаправленной сознательной деятельности.
В основе книги лежат материалы, собранные автором в течение длительной (свыше 40 лет) работы по психологическому изучению больных с локальными поражениями мозга. Большая часть этой книги посвящена, таким образом, анализу изменений, наблюдаемых в поведении таких больных.
Нейропсихология стала за последние десятилетия важной практической областью медицины, позволившей привлечь новые приемы с целью ранней и возможно более точной топической диагностики локальных поражений мозга и к научно обоснованному восстановлению функций.
Одновременно она явилась мощным импульсом к пересмотру основных представлений о внутреннем строении психологических процессов, важнейшим средством создания теории мозговых основ психической деятельности человека.
Обобщение данных, соответствующих современному этапу становления нейропсихологии, представляет собой основную задачу этой книги.
10
Глава I
ТРИ ИСТОЧНИКА ЗНАНИЙ О ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ МОЗГА
Наши знания о функциональной организации мозга животных и человека являются результатом использования трех следующих методических процедур:
во-первых, сравнительно-анатомических наблюдений;
во-вторых, физиологического метода раздражений отдельных участков мозга;
в-третьих, метода разрушения ограниченных участков мозга, а при исследовании функциональной организации мозга человека — клинических наблюдений над изменением поведения больных с локальными поражениями мозга.
Остановимся подробно на каждом из этих источников в отдельности.
1 СРАВНИТЕЛЬНО-АНАТОМИЧЕСКИЕ ДАННЫЕ
Изучение строения нервной системы — основного аппарата связи животного с внешним миром и регуляции его поведения — дает неоценимый материал для анализа того, что является субстратом психической деятельности на отдельных этапах психического развития, как осуществлялась регуляция поведения на последовательных этапах эволюции и что отличает нервную систему животных, живущих в различных условиях внешней среды и характеризующихся различными формами поведения.
Тесная связь строения нервного аппарата с уровнем организации поведения и экологическими особенностями животного позволяет широко использовать сравнительно-анатомический анализ для исследования способов жизни, особенностей поведения и основных принципов организации деятельности животных.
Рассмотрим в самом кратком виде наиболее существенное в том, что может дать сравнительно-анатомический метод для решения вопроса о мозге как органе психики.
11
Основные принципы эволюции и строения мозга как органа психики
Рассматривая строение нервной системы на последовательных этапах эволюции животного мира, можно выделить основные принципы этой эволюции.
Основной и наиболее общий принцип заключается в том, что на различных этапах эволюции отношения организма животного со средой и его поведение регулировались различными аппаратами нервной системы, и, следовательно, мозг человека является продуктом длительного исторического развития.
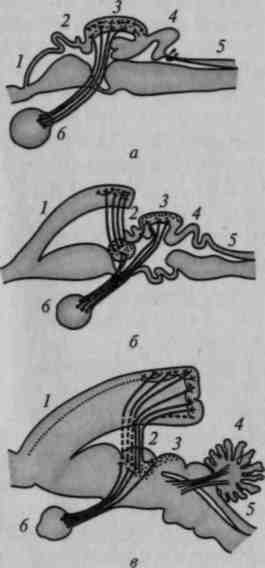
Рис. 1. Прогрессивная эволюция
соотношения аппаратов
переднего мозга:
а — мозг лягушки; б — мозг пресмыкающихся; * — мозг млекопитающих; / — передний мозг; 2 — промежуточный мозг; 3 — средний мозг; 4 — мозжечок; 5 — продолговатый мозг; 6 — глаз (по Монакову)
Хорошо известно, что на наиболее элементарных уровнях развития животного мира (например, у гидроидных полипов) прием сигналов и организация движений осуществляются диффузной, или сетевидной, нервной системой; на этом этапе эволюции единый центр, перерабатывающий информацию и регулирующий поведение животного, отсутствует и поток возбуждения определяется теми временными доминирующими очагами, которые создаются в том или ином участке нервного аппарата животного. Именно поэтому здесь можно говорить о временно доминирующих участках или органах, соответствующих этим временно наиболее возбудимым сегментам организма (Бете, 1931 и др.). В процессе эволюции диффузная сетевидная система (сохранившаяся в организме животных) уступила ведущее место новым образованиям. В передних отделах головного мозга животного концентрировались сложные рецепторные приборы, и сигналы, получаемые ими, стали направляться в передний ганглий, который перерабатывал получаемую информацию и переключал возбуждение на эфферентные пути, идущие к двигательному аппарату животного.
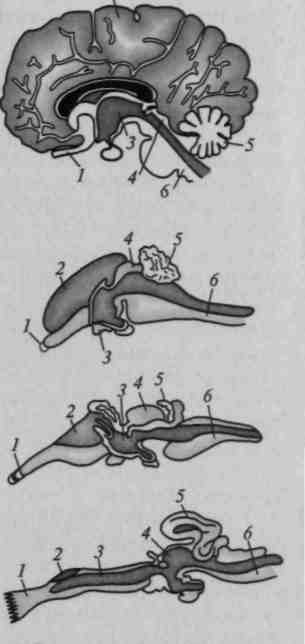
Рис. 2. Развитие объема больших полушарий головного мозга на
последовательных этапах эволюционной лестницы. Внизу — мозг акулы; выше — мозг ящерицы;
затем — мозг кролика и, наконец, вверху — мозг человека:
1 — обонятельные доли; 2 — большие
полушария; 3 ■— промежуточный мозг;
4 — средний мозг; 5 — мозжечок; 6 —
продолговатый мозг
На ранних ступенях эволюции (например, у червей) передний ганглий имел относительно простую функциональную структуру. На позднейших ступенях (например, у насекомых) по мере дифференциации системы рецепторов передний ганглий приобретает все более сложную функциональную организацию: в нем выделяются нейроны, изолированно реагирующие на обонятельные, зрительные и хемо-тактильные раздражения, промежуточные, ассоциативные нейроны и нейроны с двигательными функциями. Передний ганглий насекомых (например, пчел) становится идеальным органом реализации врожденного (инстинктивного) поведения, которое может пускаться в ход элементарными стимулами и тем не менее иметь удивительную по своей сложности программу. Эти механизмы, получившие название IRM {innate releasing mechanisms), были хорошо изучены этологами (Лоренц, 1950; , Торп, 1956; Тинберген, 1957), и мы не будем останавливаться на них.
Нервные аппараты переднего ганглия, хорошо приспособленные для реализации врожденных программ поведения, не могут, однако, обеспечить приспособления к резко меняющимся условиям среды. В таких случаях сохранение вида оказывается возможным либо благодаря избыточному производству индивидуальных особей, из которых выживают лишь очень немногие, либо благодаря выработке индивидуально-изменчивого поведения.
13
По второй линии идет развитие позвоночных. Если у низших позвоночных еще остается старый принцип сохранения индивида и вида, который хорош для существования в условиях однородной водной среды, то при переходе к наземному существованию появляется необходимость в нервных аппаратах, которые в отличие от переднего ганглия обеспечивали бы максимальную индивидуальную изменчивость поведения, соответствующую большой изменчивости условий жизни на земле.
Таким биологическим задачам отвечает головной мозг. На ранних этапах эволюции позвоночных (например, у рыб и земноводных) он допускает лишь относительно небольшую изменчивость поведения. Преобладающие формы поведения реализуются здесь аппаратами элементарного обонятельного и среднего мозга (у рыб они являются единственными и поэтому ведущими нервными образованиями). С дальнейшим развитием к ним присоединяются нервные аппараты, позволяющие животному осуществить более сложные формы анализа и приспособления к условиям среды; у птиц ведущее место занимают уже аппараты межуточного мозга (зрительный бугор, подкорковые двигательные узлы), которые образуют таламо-стриальную систему, обеспечивающую более высокий уровень поведения, названный Н.А.Бернштейном (1947) уровнем синергий.
У млекопитающих аппараты таламо-стриальной системы уступают ведущее место более сложным нервным аппаратам коры головного мозга, лежащим в основе огромного разнообразия форм индивидуально-изменчивого поведения.
Корковые аппараты в полной мере способны получать и анализировать информацию, поступающую из внешней среды, перерабатывать ее, формировать новые связи и хранить их следы. Они в состоянии заменять врожденные программы поведения сложными индивидуально-изменчивыми, обеспечивая не только выработку условных рефлексов, но и формирование значительно более сложных программ индивидуального поведения.
По мере эволюции высших позвоночных значение этих аппаратов все более возрастает, и на этапе человека, когда к естественным условиям среды прибавляются условия общественно-исторические и когда возникает язык — уникальная для человека система кодов, эти аппараты достигают такого уровня развития, что оказываются в состоянии обеспечить формы поведения, по степени сложности не имеющие равных в животном мире.
Исследователи неоднократно пытались продемонстрировать прогрессивное развитие мозга на последовательных ступенях эволюции, принимая за показатель этого развития изменение отношения массы мозга к массе тела. Пожалуй, наиболее отчетливо это увеличение описывается индексом Хауга (1958):
14
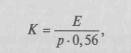
где Е — масса мозга, р — масса тела, а 0,56 — эмпирически найденный индекс; другими используется индекс Я.Я. Рогинского:
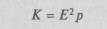
(обозначения те же).
В таблице 1 приводятся полученные с помощью этих индексов данные, показывающие, как изменялся мозг на отдельных ступенях эволюции.
Таблица 1
Увеличение относительной массы мозга на последовательных этапах филогенеза
| Индекс поХаугу | Индекс поЯ.Я.Рогинскому |
| Морская свинка 0,06 | Полуобезьяна 0,13—1,37 |
| Кролик 0,10 | Низшие обезьяны 0,56—2,22 |
| Макака 0,43 | Человекообразная обезьяна 2,03—7,35 |
| Шимпанзе 0,52 | Дельфин 6,72 |
| Человек 1,0 | Слон 9,62 |
| Человек 32,0 |
Уже эти цифры показывают, насколько большое место занимает мозг человека в системе его тела, а следовательно, и в организации его поведения.
Постепенное возрастание роли больших полушарий и коры мозга на последовательных этапах филогенеза видно уже из приведенных выше рисунков 1 и 2. Оно отчетливо подтверждается данными, представленными в таблице 2, составленной в Московском институте мозга. Цифры показывают, что если на каждое волокно зрительного нерва у крысы приходится лишь 10 нервных клеток коры, то у макаки их число увеличивается до 145, а у человека — до 500; аналогичное увеличение числа нейронов коры, приходящихся на одно нервное волокно, отмечается и в слуховой сфере (у обезьяны — преимущественно зрительного животного — увеличение не столь заметно); тот же принцип сохраняется и в отношении соответствующих отделов подкорки.
15
Таблица 2
Отношение количества нейронов к одному нервному волокну
в отдельных образованиях мозга на последовательных
ступенях эволюции, %
| Вид | Кора | Подкорковые образования | ||
| зрительная | слуховая | зрительные | слуховые | |
| Крыса | 10 | 280 | 60 | 60 |
| Макака | 145 | 300 | 145 | 20 |
| Человек | 500 | 900 | 500 | 150 |
Мы видим, таким образом, что в процессе эволюции удельный вес коры — по сравнению с нижележащими подкорковыми образованиями — непрерывно возрастает.
Естественно, что большие полушария головного мозга и его кора становятся у человека важнейшим аппаратом регуляции поведения. Существенным является тот факт, что это огромное увеличение объема и массы мозга связано не с ростом наиболее древних, стволовых, отделов мозга, а в первую очередь с развитием больших полушарий и их наиболее существенной части — коры (рис. 3).
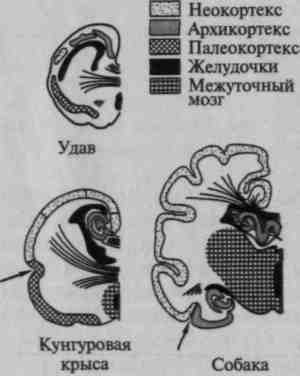
Рис. 3. Соотношение древних
и новых образований мозга
на последовательных этапах
эволюции (по Шпатиу)
Было бы неправильным думать, что все области коры человеческого мозга развиваются в процессе эволюции равномерно. Внимательный анализ показал, что развитие больших полушарий связано прежде всего с ростом новых областей коры, которые у низших млекопитающих едва намечены, а у человека составляют основную часть коры (табл. 3); древние области коры — палеокортекс (включающий образования коры, еще не отделенные от подкорковых образований), архикортекс (образования двуслойной древней коры, входящей в систему обонятельного мозга) и межуточная кора (образования, носящие переходный характер между только что упомянутыми), напротив, у человека составляют лишь незначительную часть коры, в то время как у низших млекопитающих они доминируют.
Таблица 3
Относительное изменение различных формаций коры головного мозга на последовательных этапах филогенеза, % (по И. Н. Филимонову, 1949)
| Вид | Неокортекс | Палеокортекс | Архикортекс | Межуточная кора |
| Еж | 32,4 | 29,8 | 20,2 | 17,6 |
| Кролик | 56,0 | 14,0 | 23,8 | 6,2 |
| Низшая обезьяна | 85,3 | 2,8 | 8,7 | 3,2 |
| Шимпанзе | 93,8 | 1,3 | 3,3 | 2,1 |
| Человек | 95,9 | 0,6 | 2,2 | 2,3 |
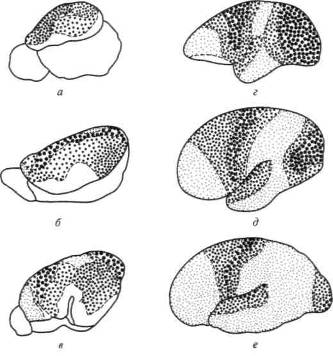
Рис. 4. Изменение в соотношении первичных, вторичных и третичных зон коры на последовательных этапах эволюции:
а — мозг ежа; б — мозг крысы; в — мозг собаки; г — мозг низшей обезьяны;
д — мозг высшей обезьяны; е — мозг человека. Крупными точками обозначены
первичные (центральные) поля ядерных зон. Точками средней величины —
вторичные (периферические) поля ядерных зон. Мелкими точками —
третичные поля (зоны перекрытия) (по Г.И.Полякову)
С переходом от высших млекопитающих (обезьян) к человеку эволюция мозга связана преимущественно с увеличением площади наиболее сложных (третичных) зон коры; площадь более элементарных отделов коры (первичных и вторичных) практически не увеличивается (а иногда даже становится меньше) (рис. 4).
В таблице 4 мы приводим данные отечественных исследователей, объединенные Московским институтом мозга.
Эти данные убедительно показывают принципиальные изменения в соотношении отдельных зон мозговой коры при переходе от низших обезьян к высшим, а затем к человеку. Они позволяют видеть, что относительный размер более просто организованной лимбической области снижается с переходом к человеку; относительный размер прецентральной (двигательной) коры остается без изменений; размер первичной (проекционной) затылочной коры у человека даже уменьшается по сравнению с таковой у обезьяны, в жизни которой зрительное восприятие играет особенно большую роль. Напротив, размеры височной области у человека значительно увеличиваются, а размеры третичных полей коры — нижнетеменной и лобной областей — возрастают в несколько раз.
Таблица 4
Относительное изменение поверхностей отдельных зон коры к величине всей коры у высших млекопитающих и человека, %
{по С.М.Блинкову, 1955; И.А.Станкевич, 1955; И.Н.Филимонову, 1949; Е.П.Кононовой, 1962; и др.)
| ^v. Область ^\коры Вид ^ч^ | Лимби-ческая | Прецент- ральная | Затылочная | Височная | Нижнетеменная | Лобная |
| Мартышка | 4,2 | 8,3 | 17,0 | 17,0 | 0,4 | 12,4* |
| Шимпанзе | 3,1 | 7,6 | 21,5 | 18,6 | 2,6 | 14,5 |
| и орангу- | ||||||
| танг | ||||||
| Человек | 2,1 | 8,4 | 12,0 | 23,0 | 7,7 | 24,4 |
* Данные о размерах лобных долей вычислены применительно к макаке.
Мы видим, таким образом, какое огромное место в коре мозга человека отводится аппаратам, связанным с приемом, переработкой (кодированием) и синтезом информации, получаемой от различных анализаторов, и аппаратам, принимающим участие в выработке и сохранении сложнейших программ поведения и контроля психической деятельности.
На детальном анализе роли этих аппаратов в структуре психических процессов человека мы остановимся далее.
18
Было бы, однако, глубоко неправильным думать, что если у человека кора головного мозга приобретает ведущую роль, то все нервные образования, которые на более низких этапах эволюции были единственными аппаратами, обеспечивающими организацию поведения, теперь совершенно отстраняются от работы.
Важнейший принцип работы мозга заключается в том, что прежние нервные аппараты сохраняются в нем, но сохраняются, если пользоваться выражением Гегеля, в снятом виде, иначе говоря, сохраняются, уступая ведущее место новым образованиям и приобретая иную роль. Они все больше и больше становятся аппаратами, обеспечивающими фон поведения, принимающими активное участие в регуляции состояний организма, передавая как функции получения, переработки и хранения информации, так и функции
создания новых программ поведения и регуляции и контроля сознательной деятельности высшим аппаратам коры головного мозга (рис. 5).
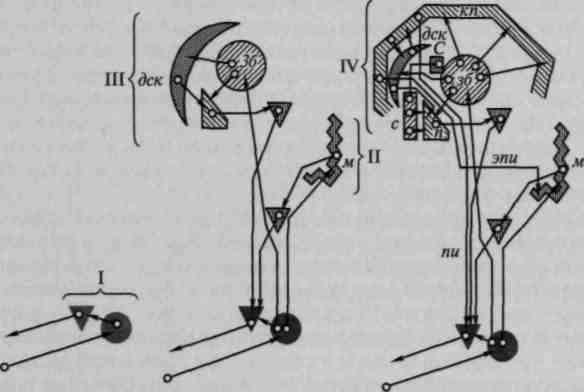
Рис. 5. Соотношение различных функциональных уровней нервной системы:
м — кора мозжечка; зб — зрительный бугор; дек — комплекс филогенетически более старых формаций коры; и — филогенетически более старая часть подкорковых узлов больших полушарий; с — филогенетически более новая часть подкорковых узлов больших полушарий; кп — наиболее поздно появляющиеся в эволюции позвоночных формации полноразвитой новой коры больших полушарий; пи — пирамидный путь для проведения к рефлекторным центрам координационного механизма формируемых в коре импульсов произвольных движений; эпи — экстрапирамидный путь для проведения влияния коры больших полушарий на кору мозжечка. Низшие рефлекторные центры спинного мозга и стволовой части головного мозга представлены черными кругами и треугольниками. Римские цифры соответствуют: I — координационному механизму; II — анализаторно-координационному механизму; III и IV — двум последовательным ступеням прогрессивного усложнения высших (надосевых) мозговых концов систем анализаторов (несколько видоизмененная схема Н.А.Бернштейна)
Забывать это положение и рассматривать аппараты коры головного мозга в отрыве от лежащих ниже образований означало бы допускать грубейшую ошибку. Мы хорошо знаем сейчас, что разные по сложности формы поведения даже у человека могут осуществляться с помощью различных уровней нервной системы.
Каждый физиолог и невролог хорошо знает, что такие простейшие элементы поведения, как сегментарные рефлексы (например, коленный рефлекс, элементарные защитные рефлексы), осуществляются лишь механизмами спинного мозга, и у больного, у которою ранение полностью отделило аппараты спинного мозга от более высоких уровней, эти рефлексы могут сохраняться или даже усиливаться, хотя и не улавливаются сознанием.
Физиологам известно также, что такая сложнейшая врожденная форма поведения, как регуляция обменного равновесия (го-меостаза), обеспечиваемая дыханием, пищеварением и терморегуляцией, осуществляется посредством механизмов, заложенных в верхних отделах ствола (продолговатом мозге, гипоталамусе); при нарушении их соответствующие процессы расстраиваются; грубые поражения этих механизмов могут привести к нарушению «витальных функций» и смерти.
Кроме того, физиологи и неврологи знают, что еще более сложные формы поведения, предполагающие обеспечение тонуса, синергий и координацию, тесно связаны с работой межуточного мозга и подкорковых двигательных узлов (таламо-спинальной системы); поражение их, не вызывая нарушения сложных познавательных процессов, приводит к грубому нарушению «фонового» поведения. Особый интерес в связи с этой проблемой представляют результаты наблюдений над больными, страдающими паркинсонизмом, накопившиеся за последние три десятилетия в результате широкого изучения эпидемического энцефалита и распространения стереотактических операций.
Наконец, хорошо известно, что наиболее сложные формы деятельности не могут быть обеспечены без участия коры головного мозга, являющейся органом высших форм поведения животных и сознательного поведения человека.
Таким образом, ясно, что сложные рефлекторные процессы и сложные формы поведения могут осуществляться разными уровнями нервной системы, каждый из которых вносит в функциональную организацию поведения свой вклад.
Последние десятилетия позволили во многом уточнить только что обозначенное положение. Было показано, что низшие уровни нервного аппарата участвуют в организации работы коры больших полушарий, регулируя и обеспечивая ее тонус.
20
Роль нижних отделов ствола и образований межуточного мозга в обеспечении и регуляции тонуса коры была показана сравнительно недавно благодаря классическим работам Мэгуна и Моруцци (1949 и др.), посвященным так называемой восходящей активирующей ретикулярной формации.
Забывать об участии низших уровней мозгового аппарата в наиболее сложных формах поведения и игнорировать тот факт, что они обеспечивают нужное состояние коры и выступают как регулятор общего фона психической деятельности, значило бы сейчас допускать серьезную ошибку. Далее (ч. первая, гл. III) мы остановимся на этом вопросе подробнее.
Аппараты стволового уровня не работают в полной изоляции от коры головного мозга и сами испытывают ее регулирующее влияние.
Работы Мак-Кэллока и др. (1946), Френча и др. (1955), Линдсли (1955, 1956, 1961), Жувэ и др. (1956, 1961), Эрнандес-Пеона (1966, 1969) и большое число исследований, на которых мы еще остановимся далее, показали огромную роль нисходящей активирующей ретикулярной формации, направляющей импульсы от коры головного мозга к нижележащим образованиям и приводящей аппараты регуляции тонуса в соответствие с информацией, получаемой субъектом, и с задачами, которые он ставит перед собой.
Данные, полученные в современных анатомических и физиологических исследованиях, позволяют сформулировать принцип вертикального строения функциональных систем мозга, иначе говоря, принцип, согласно которому каждая форма поведения обеспечивается совместной работой разных уровней нервного аппарата, связанных друг с другом как восходящими (летальными), так и нисходящими (фугальными) связями, превращающими мозг в саморегулирующуюся систему.
Этот прочно вошедший в науку принцип утверждает, что кора головного мозга, находящаяся в постоянном взаимодействии с нижележащими образованиями, не является единственным мозговым субстратом психических процессов.
Этот принцип делает понятным и те факты, которые ставили в тупик многих исследователей прошлого. Было показано, что разобщение отдельных зон коры путем круговой изоляции может не влечь за собой существенных изменений в поведении животных, в то время как подрезка коры, изолирующая ее от нижележащих образований, неизбежно приводит к значительным нарушениям ее регулирующих функций (Чоу, 1954; Сперри, 1959; Прибрам, Блейрт, Спинелли, 1966). Все это означает, что отдельные участки коры головного мозга соединяются между собой не только с помощью горизонтальных (транскортикальных) связей, но и через нижележащие образования, иначе говоря, посредством системы вертикальных связей.
21
Имея в виду сформулированные выше положения об эволюции нервных аппаратов, основных уровнях нервной системы и их взаимодействии, мы переходим сейчас к рассмотрению данных, которыми располагает сравнительная анатомия коры головного мозга.
Структурная и функциональная организация коры головного мозга
Наблюдения, показавшие, что мозг в целом и его кора в частности обладают неоднородным строением, относятся еще к началу прошлого века.
Ф. Галль, известный анатом, вошедший в историю науки как основатель фантастической «френологии» (концепции о функциональной организации мозга, исходящей из представлений о локализации сложных психических «способностей» в его ограниченных участках), впервые отличил серое вещество, составляющее мозговую кору и подкорковые серые образования, от белого вещества, состоящего из проводящих волокон, связывающих отдельные участки коры и соединяющие кору большого мозга с периферией. Однако это открытие, сделавшее Галля подлинным основателем морфологии мозга, долго оставалось без адекватной оценки, и настоящее раскрытие функций коры головного мозга, ее проводящих путей и серого вещества, заложенного в глубине больших полушарий, было сделано лишь спустя несколько поколений.
Значительный шаг вперед был сделан в 1863 г. киевским анатомом В. А. Бецом, занимавшимся микроскопическим изучением клеточного состава мозговой коры. Ему принадлежит открытие, которому было суждено стать началом целой эпохи блестящих исследований.
Описывая строение различных участков мозговой коры, он обнаружил, что их морфологическая структура в высокой степени неоднородна: если кора передней центральной извилины включает в свой состав большие, имеющие форму пирамиды нервные клетки (они получили в дальнейшем название гигантских пирамидных клеток Беца), то прилегающая к ней кора задней центральной извилины имеет совсем иное, мелкозернистое строение и совсем лишена пирамидных клеток (рис. 6).

Рис. 6. Два вида строения коры: а — передняя (моторная) кора; 6 — задняя (сенсорная) кора (по Бродману)
Позднее было установлено, что различие этих двух областей коры не только морфологическое, но и функциональное. Гигантские пирамидные клетки Беца (составляющие пятый слой коры) оказались источниками двигательных импульсов, идущих от коры к периферической мускулатуре, а передняя центральная извилина, в которой они были сосредоточены, — моторной областью коры головного мозга. Поля мозговой коры, имеющие мелкозернистое строение и отличающиеся развитым четвертым слоем нервных клеток (к их числу относятся и образования задней центральной извилины), оказались аппаратами, к которым подходят чувствительные волокна, начинающиеся в периферических органах чувств (рецепторах), а соответствующие зоны коры — первичными чувствительными образованиями коры большого мозга.
С выделением двигательных и сенсорных областей (или первичных двигательных и сенсорных центров) был сделан первый шаг к созданию функциональной карты коры головного мозга, и кажущаяся однородной масса серого вещества, покрывающая тонким слоем большие полушария, начала приобретать дифференцированный характер.
Дальнейшие сравнительно-анатомические наблюдения подтвердили плодотворность наметившегося подхода. Оказалось, что внимательное изучение «первичных» областей мозговой коры позволяет делать точные выводы о некоторых особенностях поведения животного. Следующие примеры хорошо иллюстрируют это положение.
23
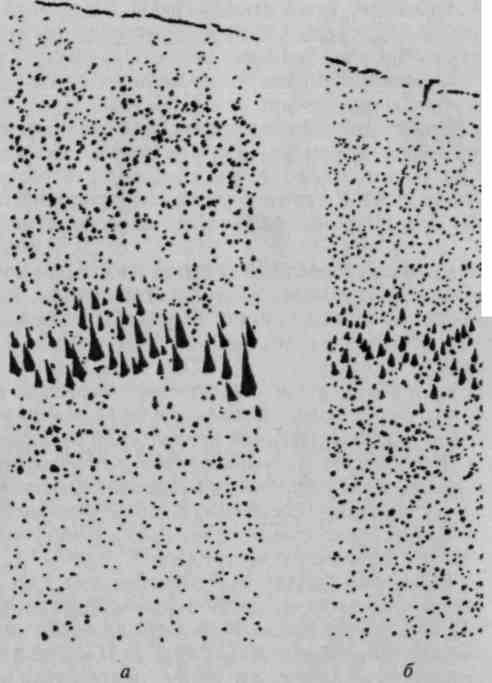
Рис. 7. Строение двигательной коры: а — австралийского медвежонка; б — летучей собаки (по Бродману)
На рисунке 7 мы приводим срезы из двигательных областей мозговой коры австралийского медвежонка (рис. 7, а) и летучей собаки (рис. 7, б). Легко увидеть, что на первом срезе присутствует сравнительно немного гигантских пирамидных клеток, в то время как на втором срезе число их значительно больше, а величина гораздо меньше. Не указывает ли нам этот факт на относительно сильные и грубые движения первого животного и более тонкие и многообразные движения второго?
К аналогичным заключениям позволяет прийти и сравнительно-анатомический анализ строения сенсорных отделов коры.
24
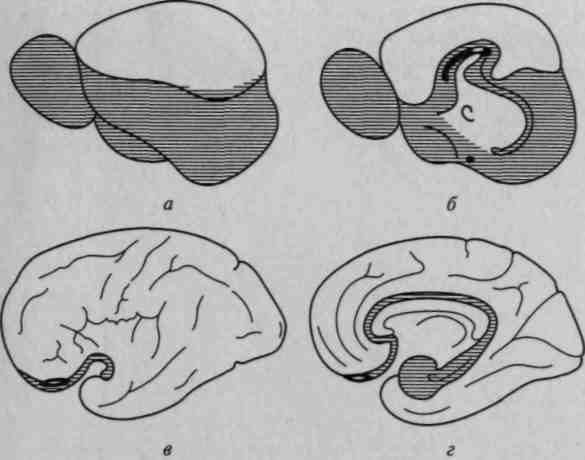
Рис. 8. Протяженность обонятельной коры:
а, б — большой мозг ежа с боковой и медиальной сторон; в, г — большой мозг
человека. Новая кора оставлена незаштрихованной. У ежа она занимает 1/4,
а у человека 11/12 всей поверхности мозга (по Экономо)
Рисунок 8 показывает то место, которое занимают образования обонятельных полей в коре головного мозга ежа (рис. 8, а, б) и человека (рис. 8, в, г). Он указывает на ведущую роль обонятельного анализатора у низших млекопитающих и незначительное место этого анализатора у человека.
Подобный вывод можно сделать и при сравнении зрительных полей животных и человека (рис. 9): значительное развитие мелкоклеточных образований зрительной коры обезьяны, составляющих у нее до 40 % площади коры (рис. 9, б), по сравнению с такими же образованиями зрительной коры крота (рис. 9, а) объясняется тем, что ведущее место в поведении первого животного занимает зрение, в то время как в поведении второго животного, ориентирующегося в окружающем мире с помощью обоняния, оно занимает лишь сравнительно небольшое место.
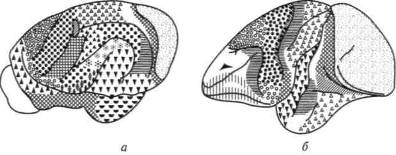
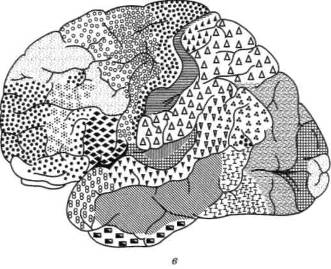
Рис. 9. Протяженность зрительной коры:
а — крота; б — обезьяны; в — человека (пунктиром обозначены мелкоклеточные образования первичной зрительной области коры)
(по Бродману)
Сравнительно-анатомическое изучение коры головного мозга, начавшееся с выделения основных, первичных, или проекционных, зон мозговой коры, существенно продвинулось за последние десятилетия.
25
Решающие успехи были сделаны еще в начале этого столетия, когда работы Кэмпбелла (1905) и Бродмана (1909), Рамон-и-Кахала (1909 — 1911) и Болтона (1933) позволили составить цитоархитектонические карты мозговой коры; эти карты, существенно уточненные Ц. и О. Фогтами (1919 — 1920) и работами Московского института мозга, позволили приблизиться к описанию основных принципов строения мозговой коры животных и человека и внесли неоценимый вклад в наши знания о мозге как органе психики.
Как показали эти исследования, новая кора головного мозга состоит из шести слоев клеток (рис. 10). Только нижние из них являются аппаратами, непосредственно связывающими мозговую кору с периферией: органами чувств (IV — афферентный слой) и мышцами (V — эфферентный слой). В IV слое коры приходят волокна, несущие импульсы, возникающие в периферических ре-
26
цепторах; именно этот слой мелкозернистых клеток особенно мощно развит в только что упомянутых первичных чувствительных зонах коры. V слой включает в себя гигантские пирамидные клетки, генерирующие импульсы к мышцам тела и дающие начало длинному двигательному пути, состоящему из нервных волокон; абсолютное преобладание этого слоя имеет место в передней центральной извилине, или двигательной зоне, коры головного мозга.

Рис. 10. Типы архитектонического строения коры больших полушарий человека:
а — тип двигательной области; б — фронтальный тип; в — париетальный тип; г — полярный тип; д — гранулярная, зернистая кора; I—V — слои коры
(по Экономо)
На рисунке 11 мы приводим схему, позволяющую проследить ход волокон от периферических органов чувств в соответствующие «проекционные» отделы коры головного мозга. Она показывает, что волокна, начинающиеся от чувствительных аппаратов кожи и мышц, прерываясь в подкорковых образованиях, приходят к коре задней центральной извилины (общечувствительная зона), а волокна, идущие от сетчатки глаза и от внутреннего уха, также переключаясь в подкорковых аппаратах, заканчиваются соответственно в затылочных и в первичных височных отделах коры.
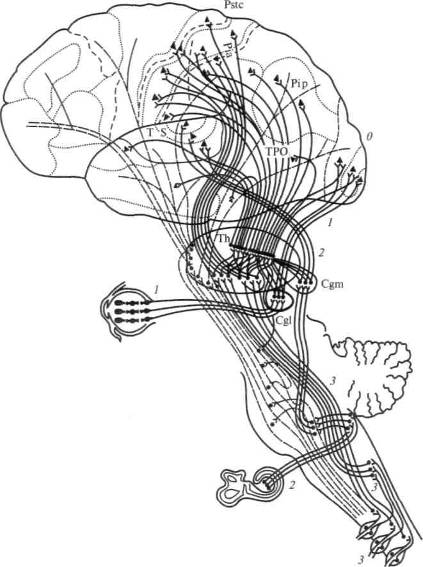
Рис. 11. Афферентные пути и чувствительные зоны коры. Жирными
линиями показаны системы анализаторов с их переключениями в
подкорковых отделах:
/ — зрительный анализатор; 2 — слуховой анализатор; 3 — кожно-кинестети-
ческий анализатор. Т — височная область; О — затылочная область; Pip —
поле 39; Pia — поле 40; Pstc — постцентральная область; ТРО — височно-
теменно-затылочная область; ТИ — зрительный бугор; Cgm — внутреннее
коленчатое тело; Cgl — наружное коленчатое тело (по Г. И. Полякову)
27
Таким образом, в коре головного мозга человека выделяются проекционная общечувствительная (теменная), зрительная (затылочная) и слуховая (височная) области.
Аналогичным образом мы можем проследить волокна, которые, начинаясь в передней центральной извилине и подходя к передним рогам спинного мозга, несут двигательные импульсы к мышцам. Эти волокна составляют двигательный, или пирамидный, путь головного мозга.
Как показали морфологические исследования, над каждой «первичной» зоной коры (с преобладающим развитием IV — афферентного или V — эфферентного слоев клеток) надстраивается система «вторичных» зон, в которых преобладающее место занимают более сложные по своему строению II и III слои. Эти слои состоят из клеток с короткими аксонами, большая часть которых или не имеет прямой связи с периферией, или получает свои импульсы из лежащих в глубине мозга подкорковых образований, осуществляющих первичную переработку приходящих с периферии импульсов. Строение этих слоев позволяет относить их уже не к простейшему — «проекционному», а к гораздо более сложному — «ассоциативному», или «интегрирующему», аппарату коры головного мозга.
Существенным для понимания функции этих слоев коры головного мозга является тот факт, что в процессе эволюции видов удельный вес их непрерывно увеличивается (рис. 12); это показывает, что процесс усложнения психической деятельности, переход от относительно простых, врожденных форм поведения животного к более сложным формам кодирования поступающей информации у человека, предполагающим сознательный характер программирования деятельности, связаны с развитием этих высших слоев мозговой коры.
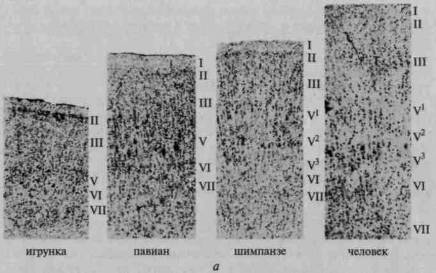
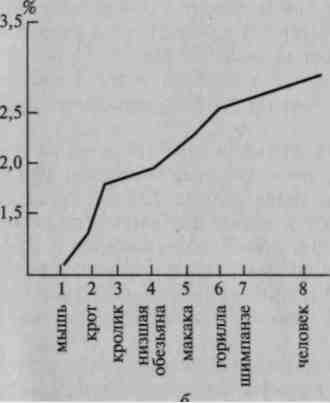
Рис. 12. Сравнительная
толщина верхних слоев коры
в филогенезе:
а — срезы; б — схема
(по данным Московского
института мозга)
Другой не менее важной функциональной характеристикой строения коры мозга животного является отношение между массой клеточных тел и массой клеточного вещества.
Исследования последнего времени показали, что в осуществлении сложных нервных процессов решающую роль играет не только тело нервной клетки, но и ее многочисленные отростки и, наконец, окружающие нейроны глиальные клетки (Хиден, 1962, 1964; Ройтбак, 1965; и др.).
Увеличение «глиального индекса» на каждой новой ступени эволюции указывает на повышение управляемости функций отдельных мозговых зон, однако лишь будущие сравнительно-анатомические исследования могут выявить его недлинный функциональный смысл.
29
Характерным поэтому является тот факт, что с эволюцией животного величина отношения глиальной ткани коры к массе ее нервных клеток все более возрастает и у человека оказывается во много раз большей, чем у млекопитающих, стоящих на более низких ступенях эволюции (табл. 5).
Таблица 5
А. Отношение массы серого вещества коры (нейронов) к массе глнальных клеток («глиальный индекс») на разных ступенях эволюции
(по Фрид, 1954)
| Вид | Слои коры | ||
| I | II | IV | |
| Мышь | 0,29 | 0,49 | 0,62 |
| Человек | 1,24-1,70 | 1,65 | 1,98 |
Б. Отношение массы серого вещества коры (нейронов) к массе глнальных
клеток («глиальный индекс») на последовательных ступенях эволюции
в пределах двигательной зоны коры
(по Банану, 1951)
| Вид | Величина клеток Беца, мк | Число клеток Беца в 1 мм3 серого вещества | Отношение массы серого вещества к массе клеток Беца, усл. ед. |
| Низшие обезьяны | 3,7 | 31,0 | 52 |
| Высшие обезьяны | — | — | 113 |
| Человек | 6,1 | 12,0 | 233 |
Аналогичная тенденция легко прослеживается в процессе созревания коры мозга человека. У плода 6 месяцев верхние слои коры едва намечены, у младенца — развиты относительно слабо, у нормального взрослого — занимают значительное место (рис. 13). В случаях врожденного слабоумия эти слои клеток недоразвиты, а у больных с органической деменцией и атрофией коры — резко сужены.
30
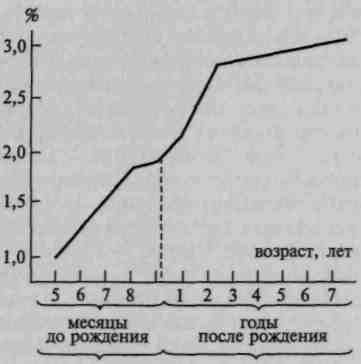
Рис. 13. Сравнительная толщина
верхних слоев коры в онтогенезе
(по Е. П. Кононовой и др.)
31
Все это указывает на то, что верхние, «ассоциативные», слои мозговой коры играют важную роль в осуществлении наиболее сложных форм психической деятельности, становление которых происходит на поздних ступенях филогенеза и на поздних этапах развития человека.
Не менее важным анатомическим фактом, позволяющим нам понять основные принципы строения мозговой коры, является неравно-
мерность распределения отдельных слоев коры в топографически различных участках коры головного мозга.
Факты показывают, что над каждой первичной областью мозговой коры, в которой преобладают низшие (афферентные или эфферентные) слои коры, надстраиваются вторичные области, в которых преобладают верхние (проекционно-ассоциационные) слои, играющие, как мы увидим далее, существенную роль в функциональной организации работы отдельных анализаторов.
Иерархическое строение мозговой коры легко можно видеть, рассмотрев топографическую карту мозга, изображенную на рисунке 14. Мы видим, что над первичными (проекционными) отделами общечувствительной коры (задняя центральная извилина) у человека надстраивается вторичная чувствительная кора, в которой преобладают верхние (проекционно-ассоциационные) слои; над первичной зрительной корой, расположенной в полюсе затылочной области, надстраивается вторичная зрительная кора, где также преобладают верхние (проекционно-ассоциационные) слои; над первичной слуховой корой, расположенной в верхних отделах височной области, надстраиваются ее вторичные отделы с тем же строением; наконец, над первичной двигательной корой, занимающей переднюю центральную извилину, — ее вторичные отделы, расположенные в премоторной области.
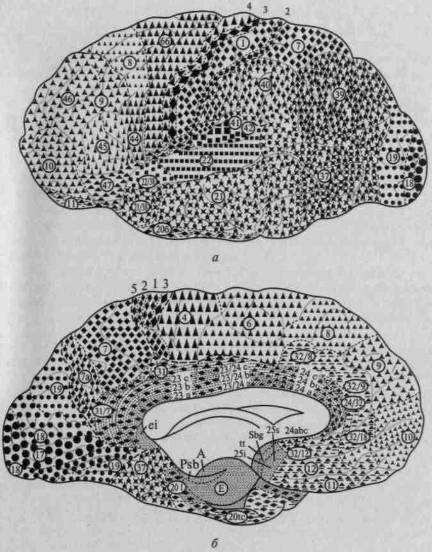
Рис. 14. Топографическая карта мозговой коры:
а — наружная поверхность; б — внутренняя поверхность. Ядерные зоны мозговой коры обозначены кружками (зрительная зона), квадратами (слуховая
зона), ромбами (общечувствительная зона), треугольниками (двигательная
зона); центральные поля выделены более крупными знаками. Зоны перекрытия
анализаторов в задних отделах полушария (теменно-височно-затылочные и
нижнетеменные отделы) обозначены смешанными знаками; в передних
отделах полушария (лобная область) — видоизмененными треугольниками;
лимбическая и инсулярная области, а также филогенетически старые зоны
коры — прерывистой штриховкой
(по Г.И.Полякову)
2 Лурия
33
Как видно из той же карты, в коре головного мозга человека можно выделить участки, которые лежат на границах между корковыми представительствами отдельных чувствительных зон мозговой коры и которые получили название третичных зон коры (или зон перекрытия коркового представительства отдельных анализаторов). Эти области коры целиком состоят из верхних (ассоциационных) слоев клеток и не имеют прямой связи с периферией. Есть все основания предполагать, что третичные зоны коры обеспечивают совместную работу корковых звеньев отдельных анализаторов, наиболее сложные интегральные функции коры головного мозга.
Как показали детальные анатомические исследования, в коре головного мозга можно выделить две группы третичных областей. Первая из них — задняя — расположена на стыке зрительной (затылочной), общечувствительной (теменной) и слуховой (височной) областей; ее с полным основанием можно обозначить как зону перекрытия корковых отделов экстероцептивных анализаторов. Вторая — передняя — расположена спереди от двигательной зоны коры и надстраивается над двигательными отделами коры головного мозга. Она связана со всеми остальными отделами коры и, как мы увидим далее, играет существенную роль в построении наиболее сложных программ поведения человека.
32
Внимательное изучение хода волокон от периферических органов чувств к коре головного мозга в полной мере подтверждает принцип иерархического строения основных отделов мозговой коры. Рисунки 14 и 15 убедительно показывают, что функциональные системы головного мозга имеют одинаковое (иерархическое) строение и что анатомические данные позволяют выделить в коре головного мозга первичные, вторичные и третичные зоны.
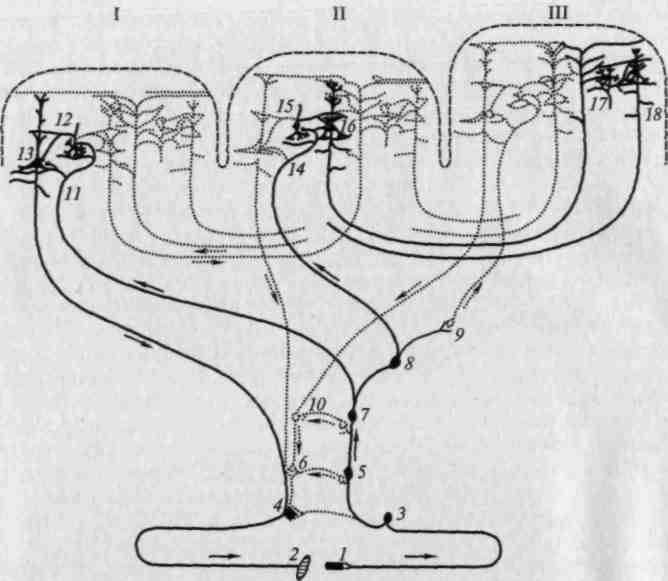
Рис. 15. Системы связей первичных, вторичных и третичных зон мозговой коры:
I — первичные (центральные) поля; II — вторичные (периферические) поля; III — третичные поля (зоны перекрытия анализаторов). Жирными линиями выделены: I — система проекционных (корково-подкорковых) связей коры; 11 — система проекционно-ассоциационных связей коры; III — система ассоциативных связей коры. 1 — рецептор; 2 — эффектор; 3 — нейрон чувствительного узла; 4 — двигательный нейрон; 5, 6 — переключательные нейроны спинного мозга и ствола; 7—10 — переключательные нейроны подкорковых образований; 11, 14 — афферентные волокна из подкорки; 13 — пирамида V слоя; 16 — пирамида подслоя III3; 18 — пирамиды подслоев III2 и Ш1; 12, 15, 17 — звездчатые клетки коры (по Г.И.Полякову)
Было бы неверным думать, что три описанных выше типа зон мозговой коры отмечаются на всех ступенях эволюции позвоночных. Факты говорят об обратном и указывают на то, что описанное нами иерархическое строение коры головного мозга является продуктом длительного исторического развития.
34
Как показывают сравнительно-анатомические данные, схематически представленные ранее на рисунке 4, в коре головного мозга ежа и крысы дифференциация первичных и вторичных зон едва намечается, а третичные зоны коры совсем отсутствуют; близкое строение имеет кора головного мозга собаки, лишь у обезьяны вторичные и третичные зоны мозговой коры отмечаются достаточно ясно. У человека иерархическое строение коры головного мозга выступает с полной отчетливостью; схема на рисунке 14 показывает, что первичные участки мозговой коры занимают у него совсем небольшое место, будучи оттесненными хорошо развитыми вторичными участками, и что третичные зоны мозговой теменно-височно-затылочной и лобной коры становятся здесь наиболее развитыми системами и занимают подавляющую часть коры больших полушарий.
В самое последнее время были получены очень важные данные, помогающие заполнить существенный пробел в наших знаниях о предыстории человеческого мозга.
Мы только что привели данные, касающиеся основных тенденций прогрессивного развития вторичных и третичных зон коры на последовательных этапах эволюции животного мира. Один вопрос остался, однако, неосвещенным: можно ли проследить такую же тенденцию и на последовательных этапах антропогенеза? Знаменуется ли переход от приматов к гоминидам и последующий переход от древних форм предчеловека к более поздним и совершенным формам соответствующим возрастанием удельного веса вторичных и третичных зон коры, отражающим усложнение условий жизни и переход от животной эволюции к социальной истории человека?
До сих пор мы знали лишь то, что объем головного мозга от высших обезьян к человеку постепенно растет и что с переходом к человеческой истории этот рост останавливается. По вычислениям Бонина (1934), средняя емкость черепа кроманьонца равнялась в среднем 1570 см3, приблизительно такая же емкость черепа характерна и для представителей неолита (1525 — 1533 см3), железного века (1514 см3) и различных периодов истории Древнего Египта (1390-1495 см3).
Означает ли это, что на всем протяжении антропогенеза соотношение отдельных образований мозговой коры оставалось неизменным?
До недавнего времени сколько-нибудь обоснованный ответ на этот вопрос казался невозможным. Только в последние годы были достигнуты первые успехи в решении этой проблемы. Связаны они с использованием оригинального метода получения слепков мозга с внутренней поверхности черепа (эндокраниума) различных предков человека. Наряду с учеными других стран большую работу в этом направлении проделала отечественный антрополог
35
В. И. Кочеткова (1966 и др.). Создавая слепки мозга с внутренней поверхности черепа ископаемого человека и пользуясь некоторыми опорными пунктами (брегма, отпечатки сильвиевой и передней центральной извилин, а также сосудов, оставляющих свой след на внутренней поверхности мозга), она смогла получить приближенные реконструкции, позволяющие определить соотноше-
36
ния отдельных частей мозга на последовательных этапах праистории (рис. 16). Дальнейший математический подсчет полученных данных позволил Кочетковой сделать некоторые выводы, имеющие самое непосредственное отношение к ответу на интересующий нас вопрос. Кратко остановимся на них.
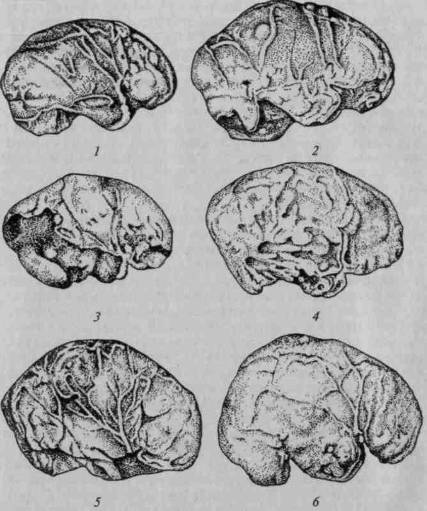
Рис. 16. Развитие мозга на последовательных этапах антропогенеза (по слепкам с эндокранов):
1 — питекантроп; 2 — синантроп; 3 — неандерталец (Брокен-Хилл); 4 — то же (Ля Шапелль); 5 — то же (Пржедмост); 6 — современный человек
(по Тильней и Дюбуа)
В древнейшей истории человека можно выделить четыре больших этапа, представителями которых являются соответственно австралопитеки (или прегоминиды), жившие 1— 4 млн лет назад, архантропы (питекантропы, синантропы), жившие 200 тыс. — 1 млн лет назад, палеоантропы (неандертальцы, люди, найденные при раскопках в Ля Шапелле, Брокен-Хилле, Тешик-Таше), жившие 200 — 400 тыс. лет назад, и неоантропы (кроманьонцы), жившие 10 — 40 тыс. лет назад.
От австралопитеков остались лишь отдельные признаки предкультуры, архантропы и палеоантропы имеют четкие признаки шелльской, ранней и поздней ашёльской и мустьерской культуры; после неоантропов встречаются более богатые находки позднего палеолита. Охотничий образ жизни, требующий четкой ориентировки в пространстве, и употребление, а затем и производство элементарных орудий знаменовали первую фазу становления человека; на следующем этапе к нему, по-видимому, присоединяется возникновение языка, и именно эти два основных признака и составляют поворотный пункт, отделяющий естественную эволюцию животных от общественной истории человека.
Как показали сравнительные исследования В. И. Кочетковой, сводные результаты которых приводятся далее, соотношения частей мозга у представителей указанных выше четырех этапов эволюции существенно различны.
На рисунке 17 мы привели сравнительную серию фронтальных срезов и сводную схему горизонтальных срезов через слепки мозга — за основу сравнения в обоих случаях берется слепок мозга высшей обезьяны (шимпанзе), который последовательно сопоставляется со слепками мозга архантропов, палеоантропов и современных людей.
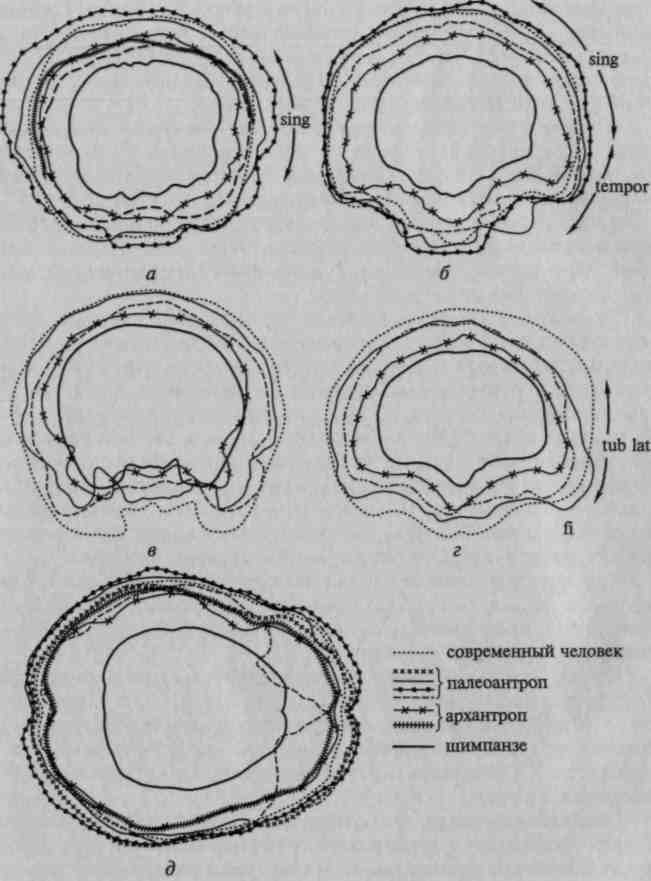
Рис. 17. Развитие отдельных областей мозга на последовательных этапах
антропогенеза:
а — фронтальный срез через нижнетеменную область (угловая борозда);
б — то же (надкраевая борозда); в — то же (височная область); г — то же
(заднелобная область); д — горизонтальный срез (по В. И. Кочетковой)
Нетрудно заметить, что слепок мозга антропоидной обезьяны имеет сферическую форму; отдельные части его не выступают 'отчетливо ни на фронтальных, ни на горизонтальных срезах.
Иное мы видим на слепках мозга ранних гоминид.
Уже на срезах через заднетеменную область (g. supramar-ginalis и g. angularis) (рис. 17, а, б) появляется выраженное местное увеличение мозга, заметное у архантропа и особенно резко выступающее у палеоантропа; аналогичную картину можно видеть на срезах через височную область (рис. 17, в), которая особенно мощно разрастается у палеоантропов. Исключительный интерес представляют срезы через заднелобную область (рис. 17, г), на анализе которых следует остановиться особо. Как показывают данные, нижнелобная область начинает существенно развиваться уже у синантропов и достигает значительного развития у палеоантропов; однако верхние отделы префронтальной области у синантропов по-
38
чти не развиты, несколько в большей степени они представлены у палеоантропов и только у современного человека получают наибольшее развитие. Подобную тенденцию можно увидеть при сопоставлении горизонтальных срезов (рис. 17, д).
Приведенные данные показывают, что основное развитие мозга в раннем антропогенезе идет не столько за счет наиболее элементарных (первичных), сколько за счет более сложных (вторичных и третичных) областей коры. Особый интерес для нас представляет последовательность развития отдельных областей больших полушарий. Оказалось, что из сложных областей коры в первую очередь начинают развиваться нижнетеменные области, значительный скачок в развитии которых отмечается уже у архантропов (синантропов и питекантропов), где они достигают существенных размеров. Есть все основания думать, что условия охотничьей жизни, требующие высокоразвитой способности ориентироваться в пространстве, составляют основу этого развития.
Во вторую очередь развиваются нижнелобные области мозга, которые достигают относительно больших размеров у палеоантропов. Это связано с возникновением и развитием звукового языка, который, как мы увидим далее, опирается на совместную работу височных и нижнелобных долей мозга.
Верхние отделы переднелобной области, как это отчетливо видно на рисунке 17, г, остаются относительно малоразвитыми на всех перечисленных выше этапах исторического развития прачеловека и интенсивно развиваются лишь у неоантропа и у современного человека. На рисунке 18 мы даем сводные данные, из которых видно, что у архантропов зона наиболее интенсивного роста расположена в нижнетеменной, у палеоантропов — в нижнелобной и у современного человека — в верхнелобной областях больших полушарий.
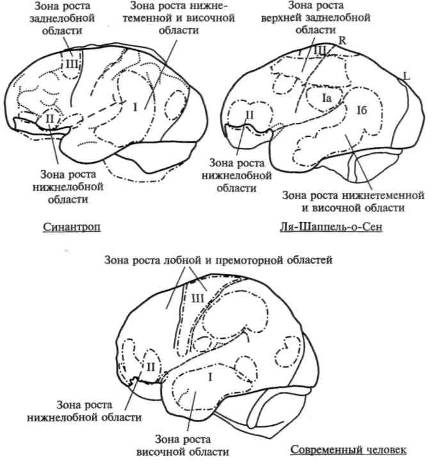
Рис. 18. Сводная схема последовательного увеличения
теменных, нижнелобных и верхнелобных отделов мозга
на последовательных этапах антропогенеза
(по В. И. Кочетковой)
Далее мы собираемся более подробно говорить об описанных здесь фактах, а также о той роли, которую играют указанные нами области больших полушарий в организации сложнейших форм сознательной деятельности. Отметим здесь только, что все приведенные данные отчетливо указывают на прямую связь эволюции головного мозга с усложнением процесса переработки и кодирования информации, с одной стороны, и с усложнением программ индивидуально-изменчивого поведения, которые отличают деятельность высших животных, — с другой.
Эти данные показывают также решающее значение эволюционного анализа анатомических изменений мозга для понимания процесса формирования сложных видов психической деятельности современного человека.
Большой интерес в плане темы данной книги представляет анализ тех изменений в мозговых структурах, которые наблюдаются в процессе развития ребенка.
39
Онтогенетические исследования основных структур человеческого мозга, начатые в свое время классиками нейроанатомии (Флексиг, 1900, 1927; Ц. и О. Фогты, 1919, 1920) и детально представленные в многолетних исследованиях Московского института мозга (Г.И.Поляков, 1938 — 1948; Е.П.Кононова, 1962; и др.), позволили получить достаточно полную картину становления основных структур головного мозга в процессе индивидуального развития. Данные этих исследований свидетельствуют о постепенной дифференциации систем мозговой коры и о неравномерном развитии отдельных мозговых структур, что, как мы убедились, имеет место также в филогенезе антропоидов.
40
Как показали исследования, ребенок появляется на свет с полностью созревшими аппаратами подкорковых образований и наиболее простых, проекционных или первичных, зон коры и с недостаточно созревшими аппаратами более сложных вторичных и третичных зон коры; это проявляется в относительно малом размере входящих в их состав клеток, в недостаточном развитии ширины их верхних слоев (имеющих, как известно, сложные ассоциативные функции), в относительно малой площади занимаемых ими территорий и, наконец, в недостаточной миелинизации их элементов.
Признаки недоразвития высших (вторичных и третичных) полей коры головного мозга исчезают постепенно и неравномерно, иными словами, коэффициенты роста отдельных полей мозговой коры неодинаковы. Если прижизненные коэффициенты роста таких относительно более простых областей коры, как лимбическая область, кора островка и кора первичных отделов центральной, затылочной и верхневисочной областей, не выходят за пределы
2,5— 5 усл. единиц, то площадь наиболее сложных, вторичных и третичных, областей коры увеличивается более интенсивно — коэффициент роста 7 —9 усл. единиц (рис. 19). Согласно материалам Московского института мозга, наиболее бурное увеличение территории последних областей отмечается в возрасте 2 — 3 лет, причем наиболее сложные лобные области созревают окончательно лишь к 6 —7-летнему возрасту (рис. 20).
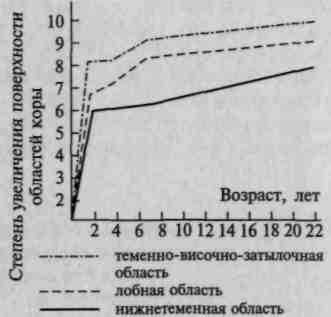
Рис. 19. Последовательное увеличение размеров первичных, вторичных и третичных зон коры в онтогенезе (по данным Московского института мозга)
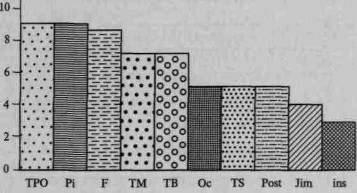
Рис. 20. Последовательное увеличение ширины различных слоев коры в онтогенезе (по данным Московского института мозга)
Таблица 6
А. Увеличение площади первичных и вторичных зон
коры от новорожденного к взрослому по отношению
к поверхности полушария, %
(по И. Н. Филимонову, 1949; С. М. Блинкову, 1955; И.А.Станкевич, 1955; Е.П.Кононовой, 1962)
| Отделы коры | Новорожденный | Взрослый | Разница |
| Затылочная | 13-14 | 12,5-13 | -0,5-1 |
| Прецентральная | 10,6 | 9,2 | -1,4 |
| Верхнетеменная | 7,3 | 7,6 | +0,3 |
| Нижнетеменная | 6,3 | 7,6 | + 1,3 |
| Височная | 16,2 | 23,4 | +7,2 |
Б. Увеличение площади вторичных и третичных зон
коры у ребенка по отношению к поверхности полушария
взрослого (по возрастам), %
| Стадия онтогенеза | Области коры | ||
| верхневисочная | нижнетеменная | теме нно- височная | |
| Новорожденный | 22 | 17,3 | 10,0 |
| 6 месяцев | 47 | 52 | 44,3 |
| 1 год | 55 | 63 | 5,4 |
| 2 года | 80 | 85 | 77,9 |
| 4 года | 88 | 88 | 73,9 |
| 7 лет | 93 | 95 | 93,7 |
| Взрослый | 100 | 100 | 100 |
42
Аналогичным образом развиваются в онтогенезе верхние (ассоциативные) слои только что упомянутых зон коры. И здесь мы отмечаем уже знакомый нам факт особенно интенсивного роста ширины этих функционально наиболее важных слоев коры к 3 — 3,5 года жизни ребенка, причем увеличение их в случае некоторых особенно сложных полей продолжается до 7- и даже 12-летнего возраста. Этот факт отчетливо говорит о том, что по мере развития ребенка возрастает роль тех видов деятельности, которые требуют совместной работы отдельных зон коры и осуществляются при ближайшем участии верхних, ассоциативных, или интегративных, ее слоев.
Более подробное описание только что указанной тенденции содержится в таблице 6, составленной на основании работ целой группы отечественных морфологов. Из таблицы видно, что если для наиболее простых (первичных, или проекционных) зон коры характерно незначительное развитие в онтогенезе, то более сложные — вторичные и третичные — зоны коры развиваются в онтогенезе весьма интенсивно.
Увеличение площади вторичных и третичных зон коры на сравнительно поздних этапах филогенеза является далеко не единственным признаком, позволяющим судить о их готовности к участию в регуляции поведения человека. Столь же существенной характеристикой является степень миелинизации соответствующих нервных образований.
Как показали классические исследования Флексига (1900, 1927), процесс миелинизации — по завершении которого нервные элементы становятся готовыми к нормальному функционированию — протекает в разных зонах коры неравномерно: если миелинизация элементов первичных (проекционных) зон заканчивается довольно рано, то процесс миелинизации во вторичных и третичных зонах коры затягивается на очень длительные сроки и в некоторых случаях продолжается до 7 —12-летнего возраста. Достаточно посмотреть на миелогенетическую карту, составленную Фогтами (рис. 21), на которой изображены зоны, элементы которых наиболее рано кончают миелинизацию (крупные кружки), зоны, которые миелинизируются в последнюю очередь (мелкие точки), и зоны, занимающие промежуточное место (средние точки), чтобы отчетливо увидеть, что аппараты, соответствующие наиболее сложным, комплексным формам психической деятельности созревают на относительно поздних этапах развития и что, следовательно, формирование психической деятельности человека идет от более простых к сложным, опосредствованным формам.
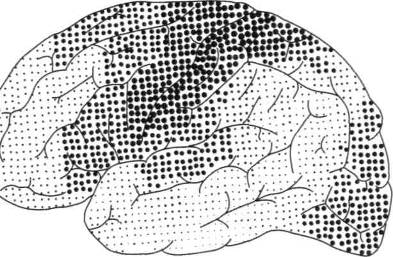
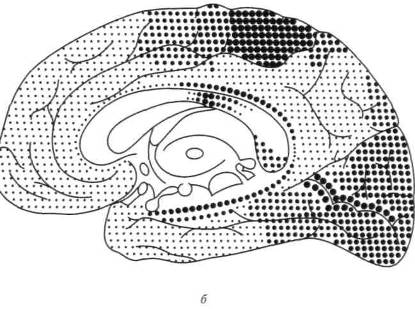
Рис. 21. Миелогенетическая карта Фогтов. Крупными точками обозначены наиболее рано созревающие отделы коры. Мелкими точками — наиболее поздно созревающие отделы коры: а — наружная поверхность; 6 — внутренняя поверхность
Далее, на протяжении книги, нам не один раз представится случай убедиться в том, насколько большое значение имеют полученные в эволюционной анатомии данные для решения важнейших проблем психологической науки.
44
2 ФИЗИОЛОГИЧЕСКИЕ ДАННЫЕ: МЕТОД РАЗДРАЖЕНИЯ
Опыты с непосредственным раздражением коры
Данные, полученные в сравнительно-анатомических исследованиях, составляют основу наших знаний о функциональной организации мозга как органа психической жизни; однако они не дают прямых указаний на нервные механизмы, связанные с приемом информации от внешнего мира и регуляцией поведения человека.
Существенный прогресс в этом направлении связан с применением метода раздражения отдельных участков мозга. Благодаря ему физиологи смогли получить сведения о непосредственной функции тех или иных мозговых систем.
Еще во второй половине XIX века исследователи раздражали отдельные пункты коры головного мозга животных, с тем чтобы выявить таким путем функции различных участков мозга. В 1871 г. были опубликованы результаты известных опытов Фрича и Гитцига, установивших, что раздражение электрическим током определенных участков коры головного мозга собаки вызывает сокращение мышц противоположных конечностей. Так была впервые выделена двигательная зона коры и положено начало точному физиологическому исследованию мозговых функций (рис. 22).
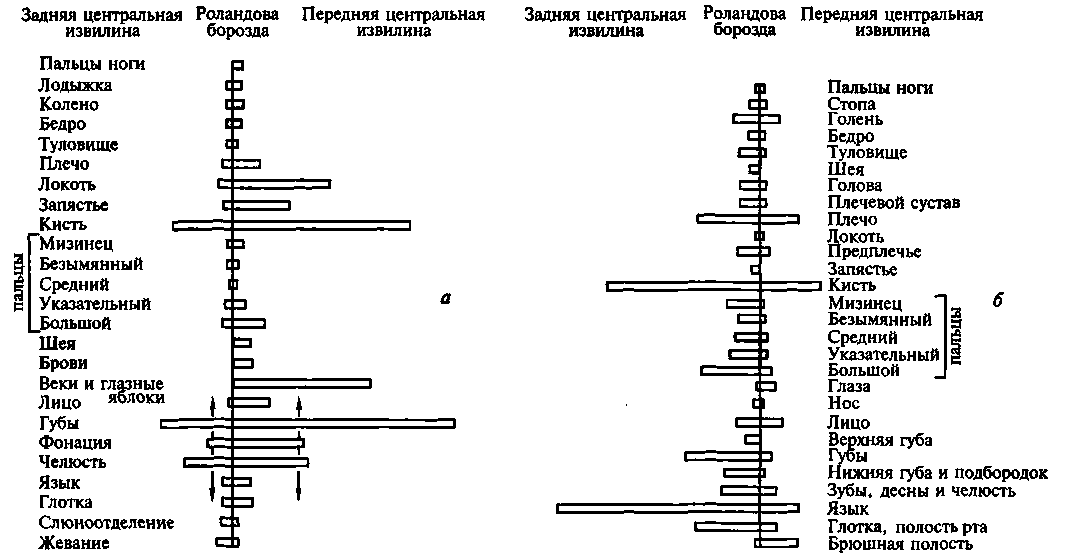
Рис. 22. Двигательные и сенсорные реакции при раздражении пре- и постцентральных отделов коры
(по Пенфилду и Расмуссену):
а — представительство двигательных процессов в мозговой коре; б — представительство сенсорных процессов в мозговой коре. Длина столбиков обозначает относительное число моторных и сенсорных реакций, вызванных раздражением точек, расположенных спереди и сзади от роландовой борозды
Спустя сравнительно небольшой период времени классические исследования, проведенные Чарльзом Шеррингтоном и его сотрудниками (Грюнбаум и Шеррингтон, 1903; Лейтон и Шеррингтон, 1917), показали, что двигательная зона коры головного мозга обезьяны имеет четкую функциональную организацию. Оказалось, что гигантские пирамидные клетки верхних отделов передней центральной извилины дают начало волокнам, идущим к нижним, а гигантские пирамидные клетки ее нижних отделов — к волокнам, идущим к верхним конечностям противоположной стороны тела. (Раздражение соответствующих пунктов двигательной зоны слабым электрическим током вызывало сокращение строго определенных мышц противоположной стороны тела.) Опыты Шеррингтона и его сотрудников положили начало изучению функциональной организации мозговой коры объективными физиологическими методами. Успехи нейрохирургии очень скоро сделали возможным дальнейший прогресс в этой области; через два десятилетия после первых публикаций Шеррингтона О. Ферстер, один из основоположников нейрохирургии, смог показать, что предложенные Шеррингтоном методические принципы пригодны и для анализа функций коры головного мозга человека. Последовавшие за исследованиями О. Ферстера работы Г. Хоффа и О.Петцля (1930) и, наконец, выдающегося канадского нейрохирурга В. Пенфилда и его сотрудников (Пенфилд и Эриксон, 1945; Пенфилд и Расмуссен, 1950; Пенфилд и Джаспер, 1959) создали прочную основу учения о функциональной организации различных зон мозговой коры человека.
45
Опыты Пенфилда не только подтвердили, что сформулированный Шеррингтоном применительно к мозгу обезьяны принцип функциональной организации моторной зоны коры верен и для мозга человека, в них были разработаны два принципиально новых аспекта исследования.
Во-первых, возможность наблюдать человека во время тонких нейрохирургических операций позволила расширить сферу изучения от двигательных функций коры до ее сенсорных функций: раздражая кору задней центральной извилины, а также затылочной и височной областей, исследователь мог спрашивать у больного об испытываемых им ощущениях и таким образом выявлять сенсорные функции этих зон коры. Таким образом, очень скоро оказалось возможным, не ограничиваясь исследованием функциональной организации двигательной сферы мозга, перейти к изучению функциональной организации афферентных, сенсорных систем.
Во-вторых, эти исследования позволили выйти за пределы простой констатации соответствия отдельных пунктов коры определенным участкам тела или, как это принято было говорить, принципа соматотопической организации первичных зон мозговой коры. Пенфилд показал, что как передняя, так и задняя центральные извилины организованы не столько по принципу линейной зависимости между размерами соответствующих участков тела и их проекциями на кору головного мозга, сколько по функциональному принципу: чем большее значение имеет та или другая функциональная система, тем более обширную территорию занимает ее проекция в первичных отделах коры 'головного мозга.
Оказалось, что проекции туловища, бедра или плеча занимают в передней и задней центральных извилинах относительно небольшую площадь; проекция руки — гораздо большую, а проекции губ и языка — еще большую площадь (рис. 23). Оказалось далее, что проекции 3-го и 4-го пальцев в коре сравнительно малы, тогда как участки, раздражая которые можно вызвать движения большого или указательного пальца, очень велики.
Рис. 23. Схема проекционных
путей первичных моторных и
сенсорных зон коры
(по Пфейфферу и Кригу)
Так возникли известные схемы Пенфилда (рис. 24), наглядно показывающие, что чем более управляемым является тот или иной орган, тем шире он представлен в мозговой коре.
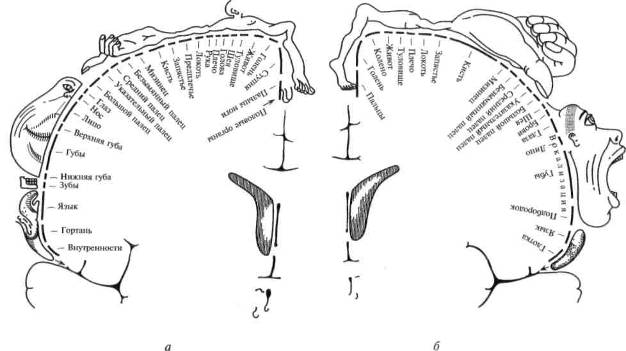
Рис. 24. Сравнительная площадь проекции отдельных поверхностей тела в проекционных зонах мозговой коры
(по Пенфилду):
а — корковая проекция чувствительности; 6 — корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения
Еще более углубили наши представления о физиологических механизмах первичных зон коры исследования, в которых было показано, что колонки клеточных элементов, соответствующие функционально наиболее важным системам органов, наиболее чувствительны к раздражениям и что существует «дифференциальная реактивность» (Differential accessibility) систем нервного аппарата, имеющих различное функциональное значение (Маунт-кастл, 1966; Филлипс, 1956, 1966). В этом отношении важны также многочисленные данные, показывающие, что пороги возбудимости клеточных элементов могут меняться под влиянием воздействий со стороны нижележащих отделов мозга (Мэгун и Моруцци, 1949; Линдсли и др., 1949; Френч, 1952; Джаспер, 1963, 1966; Эрнандес-Пеон, 1966; и др.).
Применение описанных физиологических приемов исследования функциональной организации мозга не ограничилось опытами с раздражением первичных зон мозговой коры. Результаты, значение которых для понимания функциональной организации коры головного мозга трудно переоценить, были получены в экспериментах со стимуляцией вторичных отделов мозговой коры, отличающихся, как было указано ранее, мощным развитием верхних (ассоциативных) слоев (рис. 25).
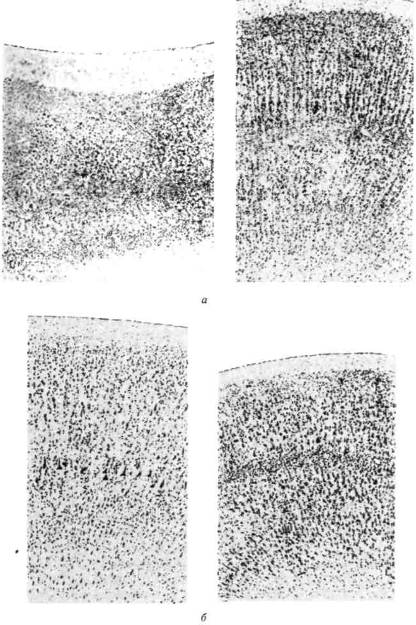
Рис. 25. Цитоархитектоническое строение первичных и вторичных зон коры (по Бродману):
а — задние отделы коры; б — передние отделы коры
Едва ли не самые интересные результаты были получены американским исследователем Мак-Кэллоком (1943, 1944), сумевшим ответить на вопрос о том, как далеко распространяется возбуждение, вызываемое раздражением определенного пункта коры головного мозга. Предложенный автором нейронографический метод был очень прост: небольшую бумажку, смоченную раствором стрихнина, налагали на определенный участок коры головного мозга, тем самым раздражая его. Затем электроды последовательно прикладывали к соседним участкам, «прощупывая» таким образом, далеко ли распространяется вызванное раздражение.
Эксперимент показал, что если раздражение первичных зон коры распространялось лишь на области, непосредственно прилегающие к раздражаемому пункту, то возбуждение, вызванное раздражением вторичных зон коры, охватывало целый комплекс зон коры, иногда расположенных на значительном расстоянии от раздражаемого пункта (рис. 26). Этот факт свидетельствует о том, что в процесс возбуждения, возникающий во вторичных отделах коры, вовлекаются большие системы нервных элементов и, таким образом, обеспечиваются значительно более сложные, интегральные, процессы по сравнению с теми, которые имеют место при возбуждении первичных зон.
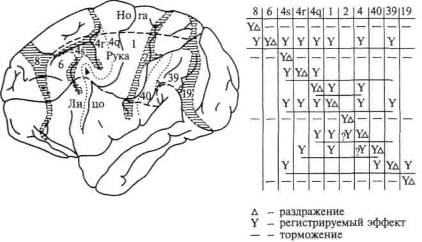
Рис. 26. Данные нейронографических опытов с раздражением первичных и вторичных (сенсорных и двигательных) отделов коры (по Мак-Кэллоку). Схема показывает, как далеко распространяется возбуждение, вызванное раздражением стрихнином определенного участка коры головного мозга шимпанзе. Поля 6, 4, 1 относятся к сенсомоторной коре. Заштрихованы так называемые депрессорные поля
50
Значение такого обширного распространения возбуждения, исходящего из вторичных зон мозга, становится ясным из опытов, в которых эффект раздражения проверялся с помощью не электрофизиологических, а психологических показателей. Результаты этих опытов настолько важны, что на них следует остановиться особо.
Эксперименты с раздражением коры головного мозга больного на операционном столе, проведенные О.Петцлем, а затем Пенфилдом и его сотрудниками, показали, что эффект раздражения определенных пунктов первичных зон коры резко отличен от того, который вызывается раздражением вторичных зон. Это с особенной отчетливостью видно при раздражениях соответствующих отделов затылочной (зрительной) и височной (слуховой) коры.
Раздражение первичных отделов коры вызывало у больного элементарные ощущения. Так, больной, у которого на операционном столе раздражали полюс затылочной зоны, заявлял, что он внезапно начинает видеть мелькающие световые точки, окрашенные шары, языки пламени и т.п., причем эти неоформленные зрительные галлюцинации (фотопсии) располагались в различных участках зрительного поля в строгой зависимости от того,
51
какое место коры раздражалось (они появлялись в верхних секторах поля зрения, если раздражались нижние участки зрительной коры, и в нижних секторах, если раздражались верхние участки). Аналогичная картина наблюдалась при раздражении первичных участков височной (слуховой) коры, с той только разницей, что в этих случаях у человека появлялись элементарные слуховые галлюцинации (шумы, тоны).
Совершенно иными явлениями сопровождается раздражение вторичных отделов коры. Раздражение передних отделов затылочной области (вторичной зрительной коры) вызывает сложные оформленные зрительные образы: испытуемый видит людей, зверей, бабочек, человека, идущего или делающего знаки рукой, птиц в полете и т. п. Следовательно, возбуждение, вызванное раздражением этой области коры, не только распространяется на более обширные площади, но и приводит к всплыванию законченных зрительных образов, сформировавшихся у человека в прежнем опыте.
Аналогичные факты отмечаются при раздражении соответствующих отделов височной (слуховой) коры. Если, как уже было указано, раздражение первичных отделов слуховой коры (трудноосуществимое из-за того, что первичные зоны слуховой коры расположены в малодоступной поперечной извилине височной области) вызывает лишь ощущение тонов или шумов, то раздражение вторичных отделов слуховой коры, расположенных в верхней и средней извилинах височной области, приводит к появлению сложных слуховых галлюцинаций — музыкальных мелодий, а иногда (если раздражается височная кора левого полушария) к звучанию слов, фраз, песен, которые испытуемый слышит, полностью сознавая отсутствие их внешнего источника, так что они оказываются чем-то средним между реальными слуховыми ощущениями и слуховыми воспоминаниями.
Интересны опыты с раздражением еще более сложных отделов височной коры. Пенфилд, который провел большую серию таких исследований, мог наблюдать, что раздражение этих областей коры вызывает сложные сценоподобные галлюцинации — целые картины, иногда включающие в свой состав как зрительные, так и звуковые компоненты. Помимо опытов с раздражением определенных пунктов коры электрическим током аналогичные явления наблюдаются в тех случаях, когда рубец, образовавшийся в мозговой ткани (или оболочке мозга) после ранения или кровоизлияния, оказывает постоянное раздражающее воздействие на соответствующие участки мозга. Накопление такого раздражения может привести к эпилептическим припадкам (например, к так называемой Джексоновской эпилепсии), причем этим припадкам предшествуют явления, носящие характер зрительных или слуховых предвестников (ауры), сложность которых зависит от локализа-
52
ции рубца. Если рубец расположен в пределах первичных зон зрительной или слуховой коры, аура носит характер элементарных зрительных или слуховых галлюцинаций; если рубец расположен в более сложных — вторичных областях коры, она может принимать характер сложных, оформленных (зрительных или слуховых) галлюцинаций. Таким образом, характер ауры, предшествующей эпилептическому припадку, служит для невропатолога указанием на местоположение рубца, являющееся «эпилептогенной зоной».
Дополнение представлений об анатомическом строении отдельных зон мозговой коры данными, полученными при раздражении ее различных участков, позволяет значительно расширить наше понимание функциональной организации коры человеческого мозга.
Обе эти группы фактов подтверждают сформулированное ранее положение о том, что отдельные системы коры головного мозга имеют иерархическое строение и что возбуждение, возникающее в периферических органах чувств, сначала приходит в первичные (проекционные) зоны, отдельные участки которых представляют собой расположенные по топографическому принципу проекции соответствующих периферических рецепторов, и лишь вслед за этим распространяется на вторичные зоны коры, которые, опираясь на аппарат верхних (ассоциативных) слоев нейронов, играют интегрирующую роль, объединяя топографические (соматотопические) проекции возникших на периферии возбуждений в сложные функционально организованные системы.
По иерархическому принципу построены и двигательные отделы коры: над проекционными отделами передней центральной извилины (моторная зона) надстраиваются вторичные отделы премоторной области, располагающие мощно развитыми верхними (ассоциативными) слоями нейронов и способные превратить отдельные двигательные импульсы в целые системы возбуждений, составляющие физиологическую основу сложных предметных движений и двигательных навыков. Подробно о функциональной организации систем двигательной коры мы будем говорить далее.
Открытие системного характера построения основных зон мозговой коры является одним из главнейших этапов формирования наших представлений о функциональной организации человеческого мозга, и мы еще много раз сможем убедиться в важности этого фундаментального принципа.
Опыты с непрямой стимуляцией коры
До сих пор мы останавливались на результатах классических исследований, основанных на непосредственном электрическом или механическом раздражении коры головного мозга.
53
Существуют и другие, гораздо более естественные методы изучения функций головного мозга, которые, однако, настолько давно известны в психологии и физиологии, что мы упомянем о них здесь лишь в самых общих чертах. Некоторые из этих методов легли в основу объективного изучения поведения животного. Животному, например, предъявляли какой-нибудь стимул (обычно связанный с инстинктивной деятельностью и являющийся для него безусловным раздражителем), после чего изучали изменения в поведении животного, которые вызывались этим стимулом. И.П.Павлов предложил метод условных рефлексов, где условные раздражители (свет, звук, прикосновение), сами по себе способные вызвать лишь элементарную ориентировочную реакцию, в сочетании с безусловными раздражителями вызывают специфические — пищевые, оборонительные или половые — реакции. Этот метод открыл новые перспективы в изучении мозговой деятельности.
Работы Павлова настолько известны, что мы не будем останавливаться на них специально. Один из приемов «непрямой» стимуляции мозговой коры заслуживает, однако, того, чтобы на нем остановиться особо. Заключается этот прием в том, что раздражение адресуется не непосредственно коре, а периферическому отделу анализатора (коже, глазу, уху), после чего в отдельных участках коры головного мозга или в подкорковых образованиях прослеживается эффект такого раздражения. Один из наиболее важных вариантов этого метода называется методом вызванных потенциалов.
Рассмотрим этот метод подробнее. Исследователи, применявшие метод раздражения в его наиболее простых формах, раздражали определенные пункты коры головного мозга электрическим током и описывали те изменения в поведении животных или человека, которые возникали как следствие такого раздражения.
Существует, однако, обратная возможность: если поставить животное или человека в условия, когда на него воздействуют определенные естественные факторы, можно выявить, какие участки коры реагируют на эти воздействия изменением электрической активности. Такой прием очень распространен в современной электрофизиологии и применяется для исследования функций различных участков коры больших полушарий.
Вариант этого метода, предложенный Даусоном (1947), получил особенно большое распространение и позволил с достаточной отчетливостью показать, как именно распространяется система импульсов, вызванных зрительными (Гранит, 1956), слуховыми (Эрнандес-Пеон, 1959, 1961) или кожными (Маунткастл, 1957, 1961, 1966; и др.) раздражителями, и детально изучить те влияния, которые оказывают нижележащие нервные образования ствола мозга на состояние мозговой коры (Эди, 1956; Эрнандес-Пеон, 1959, 1961; Линдсли, 1960, 1961).
54
Наибольший интерес для учения о функциональной организации отдельных систем мозговой коры представляют, однако, данные Эдриана (1936), показавшего, что в отдельных зонах мозговой коры сенсорная периферия представлена неравномерно и что функционально наиболее важным системам в коре соответствуют большие площади. Так, оказалось, что раздражение бедра животного вызывает изменение потенциалов лишь в незначительных по величине участках коры головного мозги животного, в то время как раздражение функционально значимых органов, играющих особенно большую роль в его жизнедеятельности (например, пятачка у свиньи, губ у овцы), ведет к появлению вызванных потенциалов в обширных территориях мозговой коры (рис. 27).
Рис. 27. Территория появления
вызванных потенциалов в коре
головного мозга свиньи
при раздражении пятачка
(по Эдриану)
Таким образом, факты показали, что функциональную карту мозговой коры можно составить с помощью не только метода непосредственного раздражения ее участков, но и метода непрямого — естественного или электрического (в частности, метода вызванных потенциалов) и что этот путь исследования также подтверждает принцип функциональной организации коры, о котором мы уже упоминали.
Метод вызванных потенциалов помогает сделать еще один существенный шаг в прослеживании более сложных форм работы основных систем головного мозга.
Как показал ряд наблюдений (Линдсли, 1961; Гарсиа-Аустт, 1963; И.А.Пеймер, 1958, 1966; Д.А.Фарбер, 1969), сложные воздействия (рассматривание сложных фигур и т.п.) вызывают в мозговой коре существенно иные электрофизиологические процессы (вызванные ответы) по сравнению с относительно более простыми. При этом меняются как латентные периоды вызванных потенциалов, так и их конфигурация.
На рисунке 28 мы приводим иллюстрацию этого положения, заимствованную из работы Э. Г. Симерницкой, проведенной в нашей лаборатории. Как показали эти опыты, у нормальных взрослых испытуемых зрительные стимулы порождают вызванные потенциалы только в затылочно-теменных отделах коры, причем эти потенциалы появляются с латентным периодом 80 — 100 мс; в отличие от этого задача дифференцировать предъявляемые зрительные стимулы по длительности приводит к появлению вызванных потенциалов не только в затылочно-теменных, но и в центрально-лобных отделах коры, причем эти потенциалы имеют значительно больший латентный период (от 120 до 260 мс) (Э. Г. Симерницкая, 1970).
Рис. 28. Изменение вызванных потенциалов (ВП) в зависимости от изменения задачи. Динамика вызванных потенциалов при изменении сигнального значения раздражителя у здоровой испытуемой. Ожидание болевого раздражителя (а, 2, 3) вызывает локальные изменения ВП в передних отделах полушарий, проявляющиеся в укорочении временных параметров ВП в передних отделах полушарий по сравнению с «фоновыми» показателями (а, 1). При отмене инструкции (а, 4) исчезает возникшее изменение ВП. Различение зрительных стимулов по длительности приводит к отчетливым изменениям ВП в затылочно-теменных областях коры (б, 2 —5) в виде резкого укорочения цикла ВП. Отмена инструкции (а, 6) снимает эффект активации. Инструкция, требующая по возможности быстрее реагировать двигательной реакцией в ответ на появление зрительного стимула, вызывает четкие изменения ВП в двигательных областях коры, показывая, что ВП могут быть в разных отношениях к моменту ответного действия, опережая его (в, 1), совпадая с ним (в, 2) и даже обнаруживаясь после его завершения (в, 3). По мере автоматизации двигательных навыков в центрально-лобных отведениях начинают регистрироваться две высокоамплитудные волны, одна из которых предшествует двигательному ответу, а другая — сопровождает его (в, 4, 5) (по Э. Г. Симерницкой)
55
Все это показывает, что метод вызванных потенциалов позволяет получать факты, убедительно говорящие о том, что более сложные формы активной психической деятельности вызывают более обширные процессы в коре головного мозга, вовлекая в совместную работу более сложные системы корковых зон.
Далее мы увидим, какое большое значение имеет это положение для понимания основных форм работы человеческого мозга как органа психической деятельности.
Опыты с анализом функций отдельных нейронов
Метод вызванных потенциалов позволил сделать еще один существенный шаг по пути познания наиболее глубоких, интимных механизмов работы головного мозга, полное значение которого будет оценено, возможно, только следующим поколением исследователей.
Развитие экспериментальной нейрофизиологии за последние десятилетия дало возможность перейти от объективного анализа функций различных систем мозговой коры к изучению функций ее составляющих элементов — отдельных нейронов.
Вживление под контролем специальных стереотактических приборов тончайших электродов в кору головного мозга или подкорковые образования и последующее отведение токов действия от отдельных нейронов при предъявлении животному различных раздражителей позволило Юнгу (1958, 1961), Хьюбелу и Визелу (1962, 1963), а также другим авторам обнаружить, что нейроны реагируют лишь на строго избирательные раздражители. Так, в зрительной коре головного мозга были обнаружены нейроны, которые реагируют пачкой импульсов только на включение или только на выключение света. Были обнаружены также нейроны, которые реагируют как на световые, так и на звуковые или кинестетические раздражители и функция которых носит, таким образом, сложный, мультимодальный характер. Наконец, были обнаружены нейроны, которые не реагируют ни на один сенсорный раздражитель и имеют, следовательно, совсем другие функции.
Аналогичные факты были получены и при исследовании нейронов височной (слуховой) коры. Так, Кацуки (1962), Эванс и др. (1965) обнаружили в слуховой коре нейроны, которые реагируют только на высокие или только на низкие тона, причем распределение этих нейронов имело топически строго упорядоченный характер. Такие же данные были получены Маунткастлом (1957, 1959, 1966) при исследовании соматосенсорной коры.
57
Подробное изучение данных нейронов дало возможность убедиться в их высочайшей специализации. Исследования обнаружили в коре (и в соответствующих подкорковых образованиях низших позвоночных) нейроны, которые реагируют только на прямые или только на плавные закругленные линии, только на острые углы или только на округлые очертания, только на движения точки от центра к периферии или только от периферии к центру и т.д. (рис. 29).
Рис. 29. Высокоспециализированные нейроны зрительных образований
золотой рыбки, реагирующие на различные по направлению движения
точки (по Кросби-Димону)
Исследование нейронов, входящих в состав вторичных зон коры, выявило существенно иную картину. С одной стороны, в этих областях были обнаружены нейроны, специфически реагирующие на более комплексные признаки, например, на наклон и толщину линии, на конфигурацию линий (Хьюбел и Визел, 1965). С другой стороны, во вторичных зонах коры было обнаружено значительное число нейронов, реагирующих на различные по модальности сигналы, а также нейроны, которые не реагируют ни
58
на какие сенсорные стимулы и, по-видимому, имеют совсем иную функцию. В самое последнее время во вторичных зонах зрительной коры кошки были выделены нейроны, реагирующие на определенное количество стимулов, причем некоторые из этих нейронов реагируют на это число как при зрительном, так и при слуховом предъявлении (Томпсон, Майерс и др., 1970). Таким образом, уже первые данные позволяют предполагать, что вторичные зоны коры включают в себя нейроны, имеющие значительно более сложные функции по сравнению с нейронами первичных зон.
Многообразие функций отдельных нейронов мозговой коры не ограничивается, однако, только что приведенными характеристиками. Юнг (1958, 1961) первым выделил в коре нейроны, которые не реагировали ни на какие сенсорные раздражители, и предположил, что они имеют какие-то иные функции.
Дальнейшие исследования, проведенные Джаспером (1963), Е.Н.Соколовым (1959), О.С.Виноградовой (1968, 1970), показали, что многие из этих нейронов, не реагируя на специальные сенсорные раздражители, активно реагируют на смену раздражителей или изменение каких-нибудь их свойств, снижая свою активность по мере привыкания и снова активизируясь при появлении изменений (рис. 30). Это заставило исследователей предположить, что они осуществляют функцию сличения новых раздражителей со следами от старых. Оказалось, что эти нейроны, получившие название нейроны внимания (attention units), распределены по мозговым образованиям очень неравномерно: если в первичных зонах зрительной коры их относительно мало, то в образованиях лимбической области, гиппокампе, хвостатом ядре, а у человека, по-видимому, и в наиболее сложных корковых формациях лобной области коры нейроны такого типа встречаются несравненно чаще.
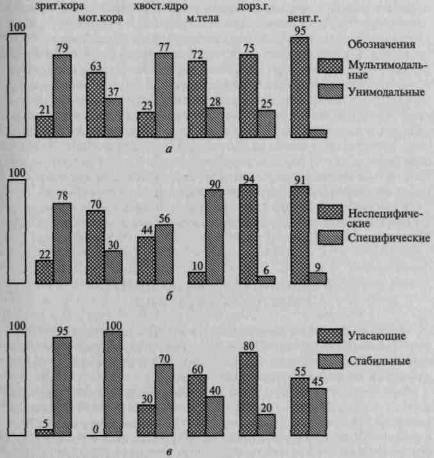
Рис. 30. Распределение нейронов различного типа в разных структурах мозга, % (по О.С.Виноградовой):
а — мультимодальные и унимодальные нейроны; б — неспецифические и специфические нейроны; в — угасающие и стабильные нейроны
Все это показывает, что в реализации активных форм психической деятельности, требующих не только получения информации, но и сличения этой информации с предшествующим опытом, принимают участие различные, в том числе далеко отстоящие друг от друга, зоны коры головного мозга и что психические процессы осуществляются сложными системами совместно работающих зон мозговой коры и нижележащих нервных образований.
Значение этого факта для понимания основных принципов функциональной организации мозга как органа психической жизни будет показано далее.
3 ФИЗИОЛОГИЧЕСКИЕ ДАННЫЕ: МЕТОД РАЗРУШЕНИЯ
Мы коротко остановились на двух источниках наших знаний о функциональной организации мозга: сравнительно-анатомических исследованиях и методах раздражения различных участков мозга непосредственным и косвенным путем.
Нам осталось ознакомиться с основными данными, полученными третьим путем, — методом выключения (или разрушения), который заключается в том, что исследователи разрушают определенные области мозга животного и прослеживают изменения в его поведении. Этому методу (или, более точно, наблюдениям над больными, у которых ранение, кровоизлияние или опухоль разрушили определенные участки мозга) суждено было сыграть основную роль в становлении новой науки о функциях мозга — нейропсихологии.
60
Еще на первых этапах физиологического анализа функции мозга было показано, что разрушение различных его участков приводит к далеко не одинаковым результатам.
Так, уже в середине XIX века было твердо установлено, что разрушение той области мозга собаки, которая содержит гигантские пирамидные клетки и, следовательно, соответствует передней центральной извилине человеческого мозга, вызывает паралич противоположных конечностей, в то время как разрушение других участков мозга (этого же полушария) не приводит к такому эффекту. Аналогичные данные были получены в опытах с высшими млекопитающими — обезьянами. Правда, эти опыты с первых же шагов выявили определенные трудности, источники которых стали понятными лишь значительно позже.
Еще в первой половине XIX века Флуранс (1842) показал, что точное ограничение двигательных функций определенными зонами мозговой коры весьма относительно: перешивая накрест нервы, идущие у петуха от определенных зон мозга к крыльям, он не наблюдал никаких изменений в двигательных функциях последних. Через некоторое время Гольц (1876—1881), разрушивший двигательные участки мозговой коры собаки, пришел к выводу, что моторные функции не ограничены у нее строго определенными участками мозга, что нарушенные функции конечностей быстро восстанавливаются после таких операций и что разрушение различных участков мозга приводит скорее к общему снижению активности поведения животного, чем к выпадению специальных, изолированных функций. Представления об отсутствии четкой функциональной организации мозговой коры у животных, существенно противоречащие всем тем фактам, которые мы приводили ранее, длительное время сохранялись в науке. Выдающийся американский исследователь К. С.Лешли (1929) описал факты, которые заставляли думать, что у крысы функции поведения связаны скорее с массой сохранившегося мозга, чем с локализацией разрушенного в опыте мозгового вещества.
Объяснить это противоречие оказалось возможным много позже, когда больше стало известно о тонком строении мозговой коры различных животных и когда были проведены сравнительные исследования, показавшие, к каким различным результатам приводит разрушение одной и той же мозговой ткани у представителей различных этапов развития животного мира.
Как уже говорилось ранее (см. рис. 9), строение мозга животных, относящихся к различным этапам эволюции, отличается различной степенью дифференцированности. Если у ежа, крота или мыши еще трудно выделить четкие, различные по своему строению, поля, сенсорные зоны коры у них еще недостаточно дифференцированы от двигательных зон, а вторичные и третичные поля почти вовсе не выделены, то на высших этапах эволюционной лестницы (например, у приматов) такая дифференцированность полей оказывается достаточно высокой; у человека она достигает высочайших пределов, причем вторичные и третичные поля доминируют во всей массе коры.
61
Уже это может служить объяснением того факта, что разрушение ограниченных зон головного мозга вызывает на различных ступенях эволюционной лестницы неодинаковый эффект и что повреждение отдельных участков мозга вызывает у низших млекопитающих (не говоря уже о низших позвоночных) менее дифференцированные дефекты, чем у высших млекопитающих и приматов (табл. 7).
Таблица 7
Последствия разрушений передних и задних отделов коры у разных представителей животного мира
| Представители животного мира | Разрушение передних (двигательных) отделов коры | Разрушение задних (сенсорных) отделов коры |
| Птица | Продолжает летать; изменений в движениях не заметно | Четко ориентируется; выбирает площадку, на которую садится |
| Собака | Движения конечностей, противоположных очагу разрушения, нарушаются; паралич противоположных конечностей, который, однако, частично претерпевает обратное развитие | Частично страдают реакции на экстероцептивные стимулы |
| Обезьяна | Стоите помощью | Значительное нарушение процессов восприятия (частично восстанавливающихся) |
| Человек | Полный и стойкий паралич конечностей, противоположных очагу разрушения | Дифференцированное и необратимое нарушение отдельных форм чувствительности |
Другая причина противоречивости результатов стала ясна из сравнительного анализа эффектов разрушения коры головного мозга. Оказалось, что разрушение коры головного мозга у птиц (у которых кора едва намечена) приводит к относительно незначительным результатам; у мыши подобное разрушение вызывает также небольшие изменения поведения; у собаки объем этих изменений возрастает, и они делаются более стойкими; у обезьяны дифференцированность и стойкость нарушений поведения, возникших в результате разрушения ограниченных участков мозга, становится несравненно более отчетливой; у человека локальные разрушения мозга вызывают стойкие нарушения различных психических процессов.
Таким образом, на последовательных ступенях эволюции поведение животного в разной степени зависит от высших отделов мозга (в частности от его коры): чем выше на эволюционной лестнице стоит животное, тем в большей степени его поведение регулируется корой и тем больше возрастает дифференцированный характер этой регуляции. Этот закон прогрессивной кортикализации функций — один из основных законов мозговой организации поведения — является ключом к пониманию причины тех противоречий, которые наблюдали отдельные авторы, изучавшие мозговую организацию поведения животных.
Роль коры головного мозга животных в организации специальных видов поведения была хорошо изучена известным американским психологом К. С.Лешли и великим русским физиологом И. П. Павловым. Следующий пример дает возможность проиллюстрировать те факты, с которыми столкнулся первый из этих исследователей, и показать трудности, связанные с решением вопроса о локализации сложных психических функций в коре головного мозга животных.
Изучая мозговую организацию зрения животных, Лешли (1930— 1942) обнаружил, что разрушение затылочной области коры приводит к тому, что обезьяна, продолжая без труда различать звуки и прикосновения, теряет возможность различать зрительно предъявляемые фигуры. Эти факты, многократно подтвержденные другими авторами (Клювер, 1927, 1937, 1941), с достаточной убедительностью показали, что задневисочные и затылочные отделы мозга этих животных имеют прямое отношение к организации зрительного восприятия и что разрушение этих областей мозговой коры неизбежно приводит к нарушению сложных форм анализа зрительных раздражений. Несмотря на всю убедительность этих данных, Лешли столкнулся с парадоксальным фактом. Оказалось, что обезьяна, лишенная зрительной (затылочной) коры, продолжала с успехом отличать треугольник от круга, успешно выполняя тем самым функции, которые, казалось бы, должны были распадаться при разрушении этого высшего аппарата зрительного восприятия.
Дальнейший анализ позволил, однако, убедиться в неправильности такого вывода: оказалось, что обезьяна, лишенная зрительной коры, различает не форму, а общую массу освещенности, различную у обеих фигур; поэтому стоило только уравнять массу света, которая отражалась обеими фигурами (уравняв площадь этих фигур), чтобы их различение стало недоступным животным с разрушенной зрительной корой.
Эти факты еще раз показывают — но на этот раз в результате применения иных методов, — что в коре головного мозга животного существуют зоны, играющие решающую роль в организации сложных форм зрительного восприятия, в то время как более простые функции (различение массы света) осуществляются элементарными механизмами подкорковых зрительных ядер.
63
Позднейшие исследования Л.Вейзкранца (1964, 1968) позволили уточнить роль сенсорных отделов мозговой коры животных в организации зрительной, слуховой и проприоцептивной информации.
Внутренние физиологические механизмы функций коры головного мозга животных по анализу и переработке сенсорной информации были с полной отчетливостью установлены работами школы И.П.Павлова, которые хронологически предшествовали всем только что упомянутым исследованиям.
Еще в первом десятилетии нашего века (XX в. — примеч. ред.) сотрудники И. П. Павлова установили фундаментальный факт: разрушение определенной области коры головного мозга собаки может привести к нарушению аналитико-синтетической работы соответствующего анализатора, не затрагивая, однако, наиболее простых форм сенсорных процессов. Так, было установлено, что собаки с удаленной височной областью коры могут реагировать на звуки, но оказываются не в состоянии выработать дифференцированные системы условных рефлексов на комплексные звуковые раздражители (Б.П.Бабкин, 1910; А.Н.Кудрина, 1910; И.Н.Кржыжановский, 1909; М. И. Эльяссон, 1908; и др.); те же закономерности были выявлены в отношении зрительного и кожного анализаторов.
Все эти исследования дали возможность убедиться в том, что кора головного мозга является аппаратом, обеспечивающим не столько элементарные сенсорные функции, сколько сложный процесс анализа и синтеза поступающей информации, и сделать тем самым шаг вперед в понимании основных физиологических законов этой сложнейшей деятельности.
Клинический материал о нарушениях психической деятельности мозга человека настолько богат и занимает такое важное место в построении нейропсихологии, что мы не будем рассматривать его здесь и посвятим его анализу специальную главу.
Хирургическое разрушение отдельных участков мозга — метод, которым в течение ряда поколений пользовались физиологи для изучения функций отдельных систем мозга, является далеко не идеальным методом исследования. Каждое хирургическое вмешательство вызывает кровоизлияние (с последующим образованием рубцов) и, таким образом, — обширные изменения мозговой ткани, что существенно затрудняет анализ роли разрушенного участка коры в построении соответствующей функции.
Вот почему в последнее время стали успешно применяться другие — бескровные — методы выключения определенных участков мозга. К числу их относится местное охлаждение отдельных участков мозга, смазывание отдельных зон мозга алюминиевой пастой (Прибрам, 1960, 1969) и, наконец, воздействие на участки мозга постоянным током (Спинелли и Прибрам, 1967), нарушающим их нормальное функционирование.
64
Преимущество этих методов перед методами разрушения состоит не только в том, что они не дают побочных, перифокальных явлений, но и в том, что такое выключение носит временный характер и что наряду с последствиями выключения может быть прослежен и процесс обратного включения соответствующих участков коры в действующие мозговые системы.
Данные, полученные благодаря применению этих методов, подтверждают и существенно обогащают те основные положения, на которых мы останавливались ранее.
Совокупность фактов, полученных в сравнительно-анатомических исследованиях, при изучении особенностей поведения, а также в физиологических работах, основанных на методе раздражения или методе выключения (разрушения) отдельных участков мозга, позволяет нам прийти к ряду решающих и в целом совершенно однозначных выводов.
Все эти источники показали, что по мере эволюции животного мира поведение все больше зависит от высших этажей мозга (его коры) или, другими словами, что на высших ступенях эволюции процесс прогрессивной кортикализации становится все более отчетливым. Кроме того, по мере эволюции функциональная организация наиболее высоких аппаратов мозговой коры становится все более дифференцированной и каждая система большого мозга приобретает отчетливую иерархическую организацию, едва намеченную у низших позвоночных, но становящуюся ведущей характеристикой мозга у приматов и особенно у человека.
Все использованные наукой источники подтверждают, что каждая из действующих систем головного мозга (зрительная, слуховая, общечувствительная и двигательная) имеет вергикильную организацию, начинаясь периферическими рецепторами, переходящими в проводящие пути, включающие наиболее простые — интегрирующие — аппараты верхнего ствола и межуточного мозга, и кончаясь высокодифференцированными аппаратами мозговой коры.
Вместе с тем источники показали, что каждая из этих систем имеет иерархическое строение; система состоит из группы надстроенных друг над другом корковых зон. В основе каждой системы лежат первичные (или проекционные) зоны коры, куда приходят импульсы, полученные посредством периферических рецепторов (органов чувств), и откуда на периферию направляются двигательные импульсы. Эти аппараты коры дробят воспринимаемую информацию на миллионы составляющих ее признаков и тем самым делают доходящие до коры возбуждения доступными кодированию и управлению.
65
Над первичными зонами надстроены вторичные зоны коры, способные благодаря преобладанию в них верхних (ассоциативных) слоев нейронов к анализу и синтезу поступающей информации, к переработке (кодированию) и хранению материала чувственного опыта и к подготовке сложных двигательных программ. Вторичные зоны коры, связанные с периферией посредством ассоциативных ядер зрительного бугра, составляют аппарат обеспечения синтетических форм работы отдельных анализаторов и занимают в коре головного мозга человека важнейшее место.
Наконец, над всем этим комплексом корковых аппаратов специализированного (модально-специфического) синтеза надстраиваются третичные зоны коры, которые выделяются в процессе эволюции позднее других и приобретают решающее значение только у человека.
Эти зоны, обладающие особенно тонким и сложным строением и располагающие мощным аппаратом ассоциативных нейронов верхних слоев коры, находятся либо на стыке вторичных отделов зрительного, слухового и общечувствительного анализаторов (вследствие чего они получили название «зон перекрытия»), либо же в передних (префронтальных) отделах большого мозга, сохраняя связь со всеми остальными отделами коры.
Изучение этих структур показывает, что они играют особенно важную роль в функциональной организации мозга, обеспечивая совместную работу отдельных анализаторов и тем самым образуя основу для получения целостной картины мира. Они являются также мозговым аппаратом, ответственным за формирование планов и программ поведения, регуляцию и контроль человеческой деятельности.
Роль третичных аппаратов мозга в человеческом поведении настолько велика, что описание их функций будет предметом рассмотрения ряда разделов этой книги.
Глава II
ЛОКАЛЬНЫЕ ПОРАЖЕНИЯ МОЗГА И ОСНОВНЫЕ ПРИНЦИПЫ ЛОКАЛИЗАЦИИ ФУНКЦИЙ
Предыдущая глава была посвящена основным источникам наших знаний о функциональной организации мозга, в ней, в частности, мы познакомились с теми данными, которые наука получила в опытах с выключением (разрушением) отдельных зон коры головного мозга животных. Однако эти данные имеют несравнимо меньшее значение для развития современных представлений о мозге как органе психики, чем результаты наблюдений над изменениями психических процессов человека при ограниченных (локальных) поражениях мозга.
Вот почему нейропсихологическое изучение больных с локальными поражениями мозга с полным основанием может считаться основным источником современных представлений о функциональной организации мозга как органа психической жизни и почему мы считаем необходимым рассмотреть получаемые этим путем факты в особой главе.
РАННИЕ РЕШЕНИЯ
Попытки рассматривать сложные психические процессы как функцию ограниченных участков мозга относятся к еще очень давнему времени. В средние века философы и натуралисты считали возможным локализовать сложные «психические способности» в трех мозговых желудочках (рис. 31). В самом начале XIX в. известный анатом Ф. Галль, впервые описавший различия между серым и белым веществом головного мозга, высказал мысль, что сложные «способности» человека интимно связаны с отдельными, строго ограниченными, участками мозга, которые, разрастаясь, образуют соответствующие выпуклости на черепе, позволяющие определять индивидуальные различия в способностях человека.
Рис. 31. Карты «трех желудочков
мозга»: 1 — воображение;
2 — мышление; 3 — память
67
Френологические карты Ф. Галля (рис. 32), представлявшие собой, по сути дела, ни на чем не основанную попытку спроецировать на мозг современную ему «психологию способностей», очень быстро были забыты. На смену им пришли попытки выделить функциональные зоны мозговой коры на основе наблюдений над изменениями в поведении человека в результате ограниченных поражений мозга.
Рис. 32. Френологические карты Ф.Галля
Клинические наблюдения больных с локальными поражениями мозга начались давно; довольно рано было установлено, что поражение двигательной зоны коры ведет к параличу противоположных конечностей, поражение заднецентральной области коры — к выпадению чувствительности на противоположной стороне тела, а поражение затылочной области мозга — к центральной слепоте.
Однако подлинное рождение учения о нарушении психических процессов с полным основанием можно отнести к 1861 году, когда молодой французский анатом П. Брока описал мозг больного, который в течение многих лет страдал грубым нарушением моторной (экспрессивной) речи; Брока установил, что в мозгу этого больного была разрушена задняя треть нижней лобной извилины. Через несколько лет дополнительные наблюдения позволили П. Брока показать, что моторная речь связана с ограниченной областью головного мозга, а именно — с задней третью нижней лобной извилины левого полушария.
Так как у всех наблюдавшихся им больных были сохранны как понимание речи, так и движения губ и языка, П. Брока предположил, что задняя треть нижней лобной извилины левого полушария является «центром моторных образов слов» и что поражение этой области приводит к своеобразному выпадению экспрессивной речи, которую он сначала назвал «афемией» и лишь потом заменил это название на существующий ныне термин афазия.
Открытие Брока имело двойной смысл. С одной стороны, впервые сложная психическая функция была локализована в определенном участке мозговой коры, причем эта локализация в отличие от фантастических попыток Ф. Галля, на поколение раньше Брока предложившего свою «френологию», носила клинически обоснованный характер.
С другой стороны, это открытие впервые показало коренное различие между функциями левого и правого полушарий мозга, выделив левое полушарие (у правшей) как ведущее, связанное со сложнейшими речевыми функциями.
68
Потребовалось лишь одно десятилетие, чтобы плодотворность открытия Брока стала очевидной: в 1873 г. немецкий психиатр К. Вернике описал случай, когда поражение другого участка (задней трети верхней височной извилины левого полушария) вызывало столь же четкую, но на этот раз обратную картину — нарушение понимания слышимой речи при относительной сохранности экспрессивной (моторной) речи. Развивая идеи П. Брока, Вернике предположил, что задняя треть первой височной извилины левого полушария является «центром сенсорных образов слова» или, как он тогда выражался, центром «понятия слова» («Wortbegriff»).
Открытие того факта, что сложнейшие формы психической деятельности могут рассматриваться как функции ограниченных участков мозга, иначе говоря, что они могут быть локализованы в ограниченных областях мозговой коры так же, как и элементарные функции (движения, чувствительность), вызвало небывалое оживление в неврологической науке, и невропатологи стали с энтузиазмом собирать факты, подтверждающие, что и другие сложные психические процессы являются функцией отдельных участков мозговой коры.
В результате такого бурного интереса к прямой локализации функций в ограниченных зонах мозговой коры в течение очень короткого срока — «блистательных 70-х годов» — в коре головного мозга были найдены «центр понятий» (в нижнетеменной области левого полушария), «центр письма» (в задних отделах средней лобной извилины этого же полушария), «центры счета», «центры чтения», «центры ориентировки в пространстве» и т.д. Позднее были описаны аппараты связей между ними, и к 80-м годам XIX в. неврологи и психиатры стали составлять «функциональные карты» мозговой коры, которые, как им казалось, окончательно разрешали вопрос о функциональном строении мозга как органа психической деятельности. Дальнейшее накопление материала благодаря наблюдениям над последствиями очаговых (локальных) поражений различных участков мозга вследствие мозговых ранений или местных кровоизлияний стимулировало эти попытки, и тенденция локализовывать сложнейшие психологические процессы в ограниченных участках мозга сохранялась в течение более чем полувека.
Данные «узких локализационистов», наблюдавших последствия ограниченных поражений мозговой коры (выпадение узнавания чисел, нарушение понимания слов или фраз, нарушение узнавания предметов, нарушение побуждений, изменения личности и т.д.), не опосредствованные тщательным психологическим анализом наблюдаемых симптомов, позволили им создать целый ряд новых гипотетических карт локализации функций в коре голов-
70
ного мозга. Наиболее подробные из таких карт были предложены немецким психиатром Клейстом (1934), который на основании обработки огромного материала наблюдений над огнестрельными ранениями мозга в течение Первой мировой войны разместил в различных участках мозговой коры такие «функции», как «схема тела», «понимание фраз», «конструктивные действия», «настроение» и т.д. (рис. 33). Принципиально такие карты немногим отличались от френологических карт Ф. Галля.
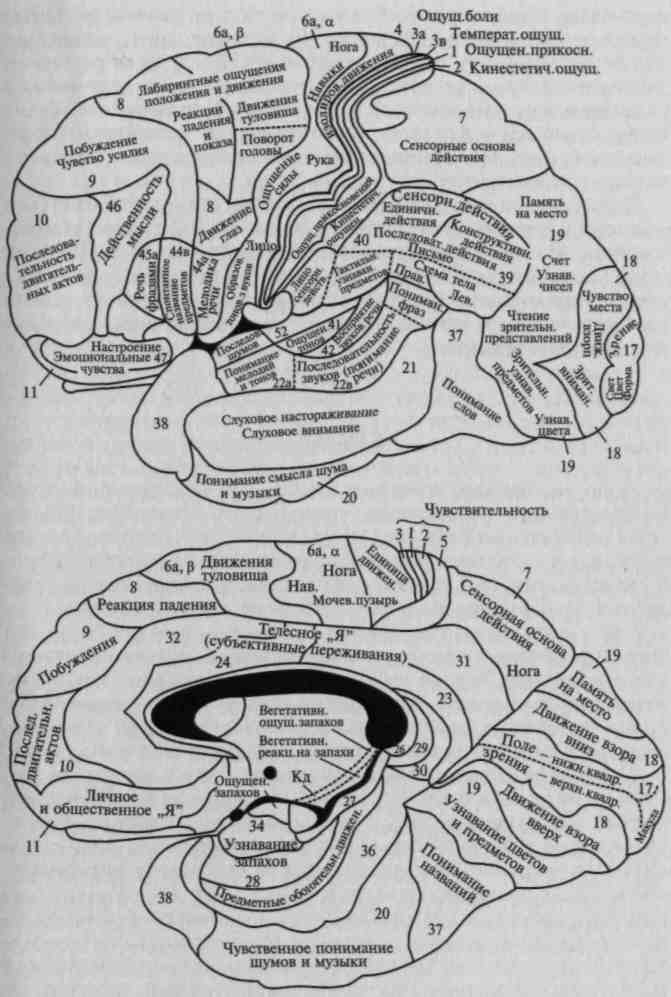
Рис. 33. Локализационная карта Клейста: вверху — наружная поверхность; внизу — внутренняя поверхность мозга
Попытки непосредственной локализации сложных психических функций в ограниченных участках мозга были настолько стойкими, что еще в 1946 г. известный американский невролог Нильсен считал возможным описывать ограниченные участки коры, которые, по его мнению, были «центрами восприятия живых объектов», отличая их от других участков, где локализовалось восприятие «неживых объектов».
2 КРИЗИС
Было бы, однако, неверным думать, что стремлениями к непосредственной локализации сложных психологических процессов в ограниченных участках мозга, или «узким локализационизмом», как его принято называть, исчерпывалось развитие неврологической мысли и что это направление не встречало противодействия со стороны достаточно влиятельных неврологов.
Уже в «блистательные 70-е годы» Брока и следовавшие за ним ученые встретили мощного оппонента в лице известного английского невролога Хьюлингса Джексона, высказавшего предположение, что к мозговой организации сложных форм психических процессов следует подходить скорее с позиций уровня их построения, чем с позиций их локализации в ограниченных участках мозга.
Положения Джексона оказались слишком сложными для его времени — лишь много лет спустя они были подхвачены и развиты выдающимися неврологами первой половины двадцатого века — Монаковым (1914), Хэдом (1926) и Гольдштейном (1927, 1934, 1948). Не отрицая того очевидного факта, что элементарные физиологические функции (такие, как кожная чувствительность, зрение, слух, движение) обеспечиваются четко очерченными участками коры головного мозга, эти исследователи выразили справедливое сомнение в том, что принцип узкой локализации приложим к мозговым механизмам сложных форм психической деятельности.
72
Указывая на сложный характер психической деятельности человека и пытаясь найти его специфические черты в осмысленном характере поведения (Монаков) или абстрактной установке и категориальном поведении (Гольдштейн), эти авторы не принимали положения о том, что соответствующие функции привязаны к ограниченным участкам мозга так же, как элементарные отправления мозговой ткани, и высказывали мысль, что сложные явления семантики или категориального поведения являются, скорее, результатом деятельности всего мозга, чем продуктом работы ограниченных участков мозговой коры. Сомнения в возможности узкой локализации сложных психических процессов приводили этих авторов либо к отрыву психических процессов от мозгового субстрата и к признанию их особой духовной природы, а именно такую позицию заняли к концу своей жизни такие выдающиеся исследователи, как Монаков (1928) и Шеррингтон (1934, 1942), либо же к попытке показать, что категориальное поведение является наиболее высоким уровнем мозговой деятельности, зависящим в большей степени от массы вовлеченного в работу мозга, чем от участия тех или иных определенных зон мозговой коры (Гольд-штейн, 1934, 1948). Таким образом, справедливые сомнения в правомерности механистического подхода «узких локализационистов» приводили либо к оживлению дуалистического признания духовной природы психических процессов, либо к оживлению идей о мозге как недифференцированном целом и решающей роли его массы в осуществлении психической деятельности, неоднократно всплывавших на протяжении всей истории изучения мозга (Флуранс, 1824; Гольц, 1876-1884; Лешли, 1929).
Легко увидеть, что если механистические представления о прямой локализации психических процессов в ограниченных участках мозга приводили исследования мозговых основ психической деятельности к тупику, то и «целостные» (или, как их иногда принято называть, «ноэтические») представления о психических процессах не создавали нужной основы для дальнейших научных исследований, сохраняя отжившие представления об обособленности духовной жизни человека и принципиальной невозможности искать ее материальную основу, равно как и столь же отжившие представления о мозге как о примитивной, недифференцированно-нервной массе.
Совершенно естественно, что такой кризис создавал необходимость поиска новых путей, которые позволили бы выявить подлинные мозговые механизмы психической деятельности человека, опираясь на те же научные принципы исследования, которые оправдали себя при изучении элементарных форм физиологических процессов, но которые были бы адекватными объекту изучения — сознательной деятельности человека, социально-исторической по своему происхождению и сложной, опосредствованной по своему строению.
Такая задача требовала коренной перестройки понимания функций, с одной стороны, и основных принципов их локализации — с другой.
73
ПЕРЕСМОТР ОСНОВНЫХ ПОНЯТИЙ
Для того чтобы подойти к вопросу о мозговой локализации сложных форм психической деятельности человека, нам нужно прежде всего остановиться на том пересмотре основных понятий, без которого правильное решение этого вопроса останется невозможным. Мы имеем в виду прежде всего пересмотр понятия «функция», затем — понятия «локализация» и, наконец, переоценку того, что называлось «симптомом», или «выпадением» функции при локальном поражении мозга.
Пересмотр понятия «функция»
Исследователи, пытавшиеся рассмотреть вопрос о локализации элементарных функций в коре головного мозга, пользуясь как методом раздражения, так и методом выключения ограниченных участков мозга, понимали «функцию» как отправление той или иной ткани.
Такое понимание, несомненно, правомерно. Совершенно естественно считать, что выделение желчи есть функция печени, а выделение инсулина — функция поджелудочной железы. Столь же правомерно рассматривать восприятие света как функцию светочувствительных элементов сетчатки глаза и связанных с нею высокоспециализированных нейронов зрительной коры, а генерацию двигательных импульсов — как функцию гигантских пирамидных клеток Беца.
Однако такое определение не исчерпывает всех аспектов понятия «функция».
Когда мы говорим о функции пищеварения или функции дыхания, понимание ее как отправление определенной ткани становится явно недостаточным.
Для осуществления акта пищеварения требуется доведение пищи до желудка, переработка пищи под влиянием желудочного сока, участие в этой переработке секретов печени, поджелудочной железы, сокращение стенок желудка и кишечника, проталкивание усваиваемого вещества по пищевому тракту и, наконец, всасывание расщепленных элементов пищи стенками тонкого кишечника.
Точно так же обстоит дело с функцией дыхания. Конечной задачей дыхания является доведение кислорода до легочных альвеол и его диффузия через стенки альвеол в кровь. Однако для реализации этой конечной цели необходимо участие сложного мышечного аппарата, в состав которого входят мышцы диафрагмы и межреберные мышцы, позволяющие расширять и сужать объем грудной клетки и управляемые сложнейшей системой нервных приборов ствола мозга и вышележащих образований.
74
Понятно, что весь этот процесс представляет собой не просто функцию, а целую функциональную систему, включающую многие звенья, расположенные на различных этажах секреторного, двигательного и нервного аппаратов.
Такая «функциональная система» (термин, введенный П.К.Анохиным, 1935, 1940, 1949, 1963, 1968; и др.) отличается не только сложностью строения, но и подвижностью входящих в ее состав частей.
Легко видеть, что исходная задача (восстановление гомеоста-за) и конечный результат (доведение питательных веществ до стенок кишечника или кислорода до альвеол) остаются во всех случаях одинаковыми (или, как иногда говорят, инвариантными). Однако способ выполнения этой задачи может сильно варьировать. Так, если основная группа работающих при дыхании мышц диафрагмы перестает действовать, в работу включаются межреберные мышцы, а если и они почему-либо страдают, включаются мышцы гортани и воздух как бы заглатывается животным. Наличие постоянной (инвариантной) задачи, осуществляемой с помощью меняющихся (вариативных) средств, позволяющих доводить процесс до постоянного (инвариантного) результата, является одной из основных особенностей работы каждой функциональной системы.
Другой особенностью является сложный состав функциональной системы, всегда включающей целый набор афферентных (настраивающих) и эфферентных (осуществляющих) компонентов.
Такое представление о функции как о целой функциональной системе резко отлично от представления о ней как об отправлении определенной ткани.
Если уж наиболее сложные вегетативные и соматические процессы построены по типу таких функциональных систем, то с еще большим основанием это понятие можно отнести к сложным функциям поведения.
Проиллюстрируем это на примере функции движения (локомо-ции), детальная структура которой была разработана замечательным отечественным психофизиологом Н.А. Бернштейном (1935, 1947, 1957, 1966).
Движение человека, имеющего намерение переместиться в пространстве, попасть в определенную точку или выполнить некоторое действие, никогда не может осуществиться одними лишь эфферентными, двигательными импульсами. Уже то, что опорный двигательный аппарат с подвижными суставами имеет, как правило, огромное число степеней свободы, умножающееся еще более от того, что в движении участвуют группы сочленений, делает движение принципиально неуправляемым одними только эфферентными импульсами. Для того чтобы начавшееся движение осуществилось, необходима постоянная коррекция его афферентными импульсами, которые сигнализируют о положении движущейся конечности в пространстве и об изменении вязкости мышц.
75
Только такое сложное строение двигательного процесса может обеспечить выполнение постоянной (инвариантной) двигательной задачи меняющимися (вариативными) средствами, т.е. достижение с помощью этих динамически меняющихся средств постоянного (инвариантного) полезного результата.
В известных опытах Хантера мышь в лабиринте достигала цели путем побежки, а когда одно из звеньев лабиринта заменялось водным бассейном, — посредством плавательных движений. В экспериментах Лешли крыса, натренированная проходить определенный путь, коренным образом меняла состав движений, когда у нее удалялся мозжечок или когда ее спинной мозг рассекался двумя половинными разрезами так, что ни одно волокно не достигало периферии, — в этих случаях, потеряв способность воспроизводить хорошо заученные движения, она достигала цели (нужного результата), двигаясь «кубарем».
Сменный характер движений, необходимых для осуществления цели, выступает также и при тщательном анализе любого двигательного акта человека: попадание в цель, например, может осуществляться различным набором движений в зависимости от исходного положения тела, другой пример — процесс письма, который можно осуществлять карандашом или ручкой, правой рукой или левой и даже ногой, не изменяя при этом ни содержания написанного, ни даже характерного для пишущего почерка (Н.А. Бернштейн, 1947).
Такое системное строение, характеризующее относительно простые поведенческие акты, в неизмеримо большей степени характерно для более сложных форм психической деятельности.
Совершенно естественно, что такие психические процессы, как восприятие и запоминание, гнозис и праксис, речь и мышление, письмо, чтение и счет, не являются изолированными и неразложимыми «способностями» и не могут рассматриваться как непосредственные функции ограниченных клеточных групп, локализованные в определенных участках мозга.
Психические процессы, как известно, формировались в течение длительного исторического развития. Будучи социальными по своему происхождению и сложными, опосредствованными по строению, они опираются на сложную систему способов и средств. Эти положения, подробно разработанные в трудах выдающегося представителя отечественной психологической науки Л.С.Выготского (1956, 1960), а также его учеников — А.Н.Леонтьева (1959), А.В.Запорожца (1960), П.Я.Гальперина (1959), Д. Б. Эльконина (1960) и др., заставляют относиться к основным формам сознательной деятельности как к сложнейшим функциональным системам и, как следствие, коренным образом пересмотреть проблему локализации их в коре головного мозга.
76
Пересмотр понятия «локализация»
Сказанное о строении функциональных систем вообще и высших психологических функций в частности заставляет нас коренным образом пересмотреть классические представления о локализации психических функций в коре головного мозга человека.
Если элементарные отправления той или иной ткани по определению четко локализованы в тех или иных клеточных группах, то о локализации сложных функциональных систем в ограниченных участках мозга или мозговой коры, конечно, не может быть и речи.
Даже такая функциональная система, как дыхание (о которой мы уже говорили), включает в свой состав настолько много элементов, что уместно вспомнить здесь слова И.П.Павлова, сказанные им при обсуждении вопроса о «дыхательном центре»: «Если с самого начала думали, что это — точка с булавочную головку в продолговатом мозгу...», то «теперь он чрезвычайно расползся, поднялся в головной мозг и спустился в спинной, и сейчас его границы точно никто не укажет...» (Поли. собр. соч. — Т. III. — С. 127). Совершенно естественно, что с локализацией высших форм психической деятельности дело обстоит еще сложнее. Высшие формы психических процессов имеют особенно сложное строение; они складываются в процессе онтогенеза, представляя собой сначала развернутые формы предметной деятельности, которые постепенно «свертываются» и приобретают характер внутренних, умственных, действий (Л.С.Выготский, 1956, 1960; П.Я.Гальперин, 1959); как правило, они опираются на ряд внешних вспомогательных средств (язык, разрядная система счисления), сформировавшихся в процессе общественной истории, опосредствуются ими и без их участия не могут быть поняты (Л.С.Выготский, 1956, 1960), они всегда связаны с отражением внешнего мира в активной деятельности и при отвлечении от этого факта теряют всякое содержание.
Вот почему высшие психические функции как сложные функциональные системы не могут быть локализованы в узких зонах мозговой коры или в изолированных клеточных группах, а должны охватывать сложные системы совместно работающих зон, каждая из которых вносит свой вклад в осуществление сложных психических процессов и которые могут располагаться в совершенно различных, иногда далеко отстоящих друг от друга участках мозга.
Едва ли не наиболее существенным в таких системных представлениях о локализации психических процессов в коре головного мозга являются два аспекта, резко отличающие работу человеческого мозга от более элементарных форм работы мозга животного.
77
Высшие формы сознательной деятельности человека всегда опираются на внешние средства (примером могут служить узелок на платке, который мы завязываем, чтобы запомнить нужное содержание, сочетания букв, которые мы записываем, для того чтобы не забыть какую-нибудь мысль, таблица умножения, которой мы пользуемся для выполнения счетных операций, и т.п.). Эти исторически сформированные средства оказываются существенными факторами установления функциональной связи между отдельными участками мозга — с их помощью те участки мозга, которые раньше работали самостоятельно, становятся звеньями единой функциональной системы. Образно выражаясь, можно сказать, что исторически сформировавшиеся средства организации поведения человека завязывают новые «узлы» в его мозговой деятельности, и именно наличие таких функциональных «узлов», или, как некоторые называют их, «новых функциональных органов» (А. Н.Леонтьев, 1959), является важнейшей чертой, отличающей функциональную организацию мозга человека от мозга животного. Именно эту сторону построения функциональных систем человеческого мозга Л.С.Выготский (1960) называл принципом «экстракортикальной» организации сложных психических функций, имея в виду под этим не совсем обычным термином то обстоятельство, что формирование высших видов сознательной деятельности человека всегда осуществляется с опорой на ряд внешних вспомогательных орудий или средств.
Другой отличительной чертой высших психических процессов человека является тот факт, что локализация их в мозговой коре не является устойчивой, постоянной, она меняется как в процессе развития ребенка, так и на последовательных этапах упражнения. Следует пояснить это положение. Известно, что каждая сложная сознательная деятельность сначала носит развернутый характер и опирается на ряд внешних опорных средств и только затем постепенно сокращается и превращается в автоматизированный двигательный навык. Так, если на первых этапах письмо опирается на припоминание графического образа каждой буквы и осуществляется цепью изолированных двигательных импульсов, каждый из которых обеспечивает выполнение лишь одного элемента графической структуры, то впоследствии в результате упражнения такая структура процесса коренным образом меняется и письмо превращается в единую «кинетическую мелодию», не требующую специального припоминания зрительного образа изолированной буквы или отдельных двигательных импульсов для выполнения каждого штриха. Аналогичным образом развиваются и другие высшие психологические процессы.
Естественно, что в процессе такого развития меняется не только функциональная структура процесса, но и его мозговая локализация: участие слуховых и зрительных зон коры, обязательное на ранних этапах формирования некоторой деятельности, перестает быть необходимым на его поздних этапах, и та же деятельность начинает опираться на иную систему совместно работающих зон (А. Р.Лурия, Э. Г. Симерницкая, Б.Тыбулевич, 1970).
78
Развитие высших психических функций в онтогенезе имеет еще одну особенность, имеющую решающее значение для функциональной организации их в коре головного мозга.
Как показал в свое время Л.С.Выготский (1960), в процессе онтогенеза меняется не только структура высших психических процессов, но и их отношение друг к другу, иначе говоря, их «межфункциональная организация». Если на первых этапах развития сложная психическая деятельность имеет более элементарную основу и зависит от основной, «базальной», функции, то на дальнейших этапах развития она не только приобретает более сложную структуру, но и начинает осуществляться при ближайшем участии более высоких по своему строению форм деятельности.
Так, если маленький ребенок мыслит, опираясь на наглядные образы восприятия и памяти, иначе говоря, мыслит припоминая, то на более поздних этапах — юношеского или зрелого возраста — отвлеченное мышление с его операциями отвлечения и обобщения развивается настолько, что даже такие относительно простые процессы, как восприятие и память, превращаются в сложные формы познавательного анализа и синтеза, и человек теперь уже воспринимает и припоминает, размышляя.
Изменение отношений между основными психологическими процессами не может оставить неизменным соотношение основных систем мозговой коры, необходимых для осуществления этих процессов. Поэтому, если в раннем возрасте поражение какой-нибудь зоны коры, обеспечивающей относительно элементарные основы психической деятельности (например, зрительных отделов коры головного мозга), неизбежно вызывает в виде вторичного, системного эффекта неразвитие более высоких, надстроенных над ней образований, то у зрелого человека, у которого сложные системы не только уже сформировались, но и стали оказывать решающее влияние на организацию более простых форм деятельности, поражение низших зон уже не имеет такого значения, которое оно имело на ранних этапах развития, и, наоборот, поражение высших зон приводит к распаду элементарных функций, приобретших сложное строение и интимно зависящих от более высокоорганизованных форм деятельности.
Сформулированное Л.С.Выготским правило, согласно которому поражение определенной области мозга в раннем детстве системно влияет на более высокие зоны коры, надстраивающиеся над ними, в то время как поражение той же области в зрелом возрасте влияет на более низкие зоны коры, которые теперь от них зависят, является одним из фундаментальных положений, внесенных в учение о динамической локализации высших
79
психических функций отечественной психологической наукой. В качестве иллюстрации его укажем, что поражение вторичных отделов зрительной коры в раннем детстве может привести к системному недоразвитию высших процессов, связанных с наглядным мышлением, в то время как поражение этих же зон в зрелом возрасте может вызвать лишь частные дефекты зрительного анализа и синтеза, оставив сохранными уже сформировавшиеся раньше более сложные формы мышления. Сказанное о системном строении высших психологических процессов заставляет нас коренным образом пересмотреть классические представления о локализации их в коре головного мозга.
Свою основную задачу теперь мы видим в том, чтобы, тщательно проанализировав совместно работающие зоны мозга, обеспечивающие выполнение сложных форм психической деятельности, выяснить место каждой из этих зон в функциональной системе и то, как меняется соотношение этих совместно работающих отделов мозга при осуществлении психической деятельности на разных этапах развития.
Естественно, что такой подход в корне изменяет и практическую форму работы психолога. Прежде чем ответить на вопрос о том, каковы мозговые основы того или иного психического процесса человека, необходимо тщательно изучить строение того психологического процесса, мозговую организацию которого мы хотим установить, и выделить в нем те звенья, которые в той или иной степени могут быть отнесены к определенным системам мозга. Только такая работа по уточнению функциональной структуры изучаемого психологического процесса с выделением его составных компонентов и с дальнейшим анализом их «размещения» по системам головного мозга позволяет подойти к новому решению старого вопроса о локализации психических функций в коре головного мозга.
Пересмотр понятия «симптом»
Классические исследования локализации психических функций в коре головного мозга, пользовавшиеся наблюдениями над изменением поведения после ограниченных (локальных) поражений мозга, исходили из упрощенных представлений, согласно которым нарушение той или иной психической функции (речи, письма, чтения, праксиса, гнозиса), наступающее в результате разрушения определенного участка головного мозга, является прямым доказательством того, что данная функция локализована в этом (теперь разрушенном) участке мозга.
80
Рассмотренные выше факты привели к коренному пересмотру этих слишком упрошенных представлений. Естественно, что нарушение общей чувствительности неизбежно указывает на поражение задней центральной извилины, равно как выпадение части зрительного поля — на поражение сетчатки, зрительных путей или зрительной коры. В этих случаях установить симптом — значит получить отчетливые данные о топической диагностике поражения и тем самым о локализации функции в нервном аппарате.
Совершенно иначе обстоит дело в случаях нарушения высших психических процессов. Если психическая деятельность является сложной функциональной системой, в осуществление которой вовлекается целый комплекс совместно работающих зон мозговой коры, то поражение каждой из этих зон может привести к распаду всей функциональной системы, и, таким образом, симптом (нарушение или выпадение той или иной функции) еще ничего не говорит о ее локализации.
Для того чтобы перейти от установления симптома к локализации соответствующей психической деятельности, нужно пройти длинный путь, важнейшим звеном которого является детальный психологический анализ структуры возникающего нарушения и выяснение тех ближайших причин, вследствие которых функциональная система распалась, или, иначе говоря, дать тщательную квалификацию наблюдаемого симптома.
Поясним нашу мысль на примере. В клинике локальных поражений мозга очень часто встречается симптом апраксии, заключающийся в том, что больной оказывается неспособным выполнить те или иные предметные действия. В классической неврологии этого было достаточно, чтобы заключить, что поражение локализуется в нижнетеменной области, считавшейся «центром сложного прак-сиса». Если апраксия носила характер затрудненного выполнения движения при отчетливо представляемой схеме, его поражение локализовалось кпереди от нижнетеменной области.
После исследований ряда физиологов (и в первую очередь выдающегося отечественного физиолога Н. А. Бернштейна) стало совершенно ясным, что всякое произвольное движение — и в еще большей мере предметное действие — представляет собой сложную функциональную систему, включающую в свой состав ряд условий, при отсутствии которых такое движение не может быть выполнено.
Для выполнения движения необходима прежде всего кинестетическая афферентация, иначе говоря, система кинестетических импульсов, которые «сообщают» мозгу о положении движущейся конечности, суставных сочленений и степени вязкости мышц. Если такие афферентные импульсы (прием и синтез которых обеспечивается общечувствительными отделами постцентральной коры) выпадают, движение теряет афферентную основу, и эффекторные импульсы, идущие от коры головного мозга к мышцам, становятся практически неуправляемыми. Как следствие, даже относительно тонкие поражения постцентральной области коры могут приве-
81
сти к своеобразной кинестетической апраксии, которая принимает характер нарушения тонких, дифференцированных движений, приводящего к невозможности обеспечить нужную позу руки, адекватную тому предметному действию, которое рука выполняет. Наличие кинестетической афферентации важное, однако недостаточное условие выполнения соответствующего движения.
Всякое движение — будь то передвижение в пространстве, попадание в цель или действие с предметом — всегда осуществляется в системе пространственных координат и необходимо нуждается в синтезе зрительно-пространственных афферентации, что обеспечивается участием третичных, теменно-затылочных, отделов коры, куда стекаются импульсы от зрительного, вестибулярного и кожно-кинестетического анализаторов.
Если эта область коры поражается и пространственный синтез распадается, с необходимостью возникает нарушение сложно построенного движения. Однако апраксия, возникающая в этих случаях, носит совсем иной характер и проявляется прежде всего в невозможности придать действующей руке нужное положение в пространстве; так, например, больной начинает испытывать затруднения, застилая кровать, и часто вместо того чтобы разместить одеяло вдоль кровати размещает его поперек, он не может сохранить правильное направление ложки, которую он держит, придавая ей часто не горизонтальное, а вертикальное положение, и т.д.
Очевидно, что такая пространственная апраксия резко отличается от описанной выше кинестетической апраксии как по структуре действия, так и по механизмам и лежащей в ее основе локализации дефекта.
Перечисленных условий, однако, все еще недостаточно для осуществления полноценного движения или действия. Каждое действие всегда состоит из цепи последовательно развертывающихся движений, каждое звено которой после реализации должно быть денервировано с тем, чтобы уступить место следующему звену. На начальных стадиях формирования эта цепь двигательных звеньев носит развернутый характер, и каждое двигательное звено требует специального изолированного импульса. С формированием двигательного навыка цепь изолированных импульсов редуцируется и сложное движение начинает осуществляться как единая «кинетическая мелодия».
Существенным является тот факт, что кинетическая организация движений обеспечивается как базальными двигательными узлами (на ранних этапах формирования элементарных двигательных синергии), так и премоторными отделами коры (на поздних этапах формирования сложных двигательных навыков). Поэтому при поражении этих отделов коры также возникает апраксия, но на этот раз она носит характер кинетической апраксии, проявляясь в невозможности
82
синтезировать двигательные звенья в одну плавную мелодию, в задержках денервации ранее выполненного звена движения и затрудненности перехода от одного двигательного звена к другому. Естественно, что структура кинетической апраксии существенно отличается от ранее описанных форм нарушения праксиса, равно как и локализация соответствующих симптомов.
Нам осталось указать на последнее условие правильности выполнения движения.
Всякое движение направлено на известную цель и реализует определенную двигательную задачу. На уровне элементарно построенного инстинктивного поведения двигательные задачи диктуются врожденными программами; на уровне сложного, прижизненно сформированного, сознательного действия они определяются намерениями, которые в свою очередь сформированы при ближайшем участии речи, регулирующей поведение человека (А. Р. Лурия, 1961). Как было показано специальными исследованиями (А. Р. Лурия, 1962, 1963, 1966, 1969; А. Р.Лурия и Е.Д.Хомская, 1966), формирование сложных намерений, осуществляющееся посредством регулирующей речи, протекает при ближайшем участии лобных долей мозга. Поэтому массивные поражения лобных долей также могут привести к апраксии, но эта апраксия целевого действия коренным образом отличается от ранее описанных форм. Как правило, она сводится к невозможности подчинить свои движения сформированному в речи намерению, к распаду сложно организованных программ и к замене осмысленного, целенаправленного действия эхопраксическим повторением движений исследующего или инертными стереотипами, теряющими осмысленный, целенаправленный характер. Мы не будем сейчас останавливаться на этом типе нарушений праксиса, поскольку далее он будет предметом специального обсуждения.
Приведенные факты позволяют нам прийти к важному заключению. Симптом нарушения праксиса (апраксия) является признаком локального поражения мозга; однако сам по себе этот симптом еще не говорит о локализации очага. Произвольное движение (праксис) является сложнейшей функциональной системой, которая базируется на сохранности ряда факторов и опирается на совместную работу целого комплекса зон мозговой коры и подкорковых образований, причем каждая из этих зон вносит свой вклад в осуществление движения. Поэтому сложное предметное движение может нарушаться при поражении различных зон коры (или подкорковых образований), однако каждый раз оно будет нарушаться по-разному.
Ближайшая задача исследователей сводится к тому, чтобы, изучив структуру наблюдаемого дефекта, квалифицировать симптом. Только такая работа, приводящая к выделению основного фактора, стоящего за наблюдаемым симптомом, позволяет прийти к заключению о локализации очага, лежащего в основе дефекта.
83
Таким образом, понятия «локализация очага» и «локализация функции» не совпадают, и для того чтобы использовать метод локальных поражений мозга для заключения о локализации функции (или, точнее, о мозговой организации функциональной системы), необходимо осуществить структурный анализ синдрома. Это и есть основной путь нейропсихологического исследования.
СИНДРОМНЫЙ АНАЛИЗ И СИСТЕМНАЯ ОРГАНИЗАЦИЯ ПСИХИЧЕСКИХ ПРОЦЕССОВ
Квалификация симптома является лишь первым этапом анализа мозговой организации психических процессов. Для того чтобы данные о локальной патологии мозга позволили прийти к надежным выводам относительно как структуры психических процессов, так и их локализации в коре головного мозга человека, необходимо перейти от квалификации единичного симптома к описанию целого симптомокомплекса, или, как это принято называть, к синдромному анализу изменений поведения, наступающих при локальном поражении мозга.
Как мы уже сказали, любая психическая деятельность человека является сложной функциональной системой, реализация которой обеспечивается целым комплексом совместно работающих аппаратов мозга, каждый из которых вносит свой вклад в обеспечение этой функциональной системы. Это практически означает, что функциональная система в целом может нарушаться при поражении большого числа зон, причем при различных по локализации поражениях она нарушается по-разному. Последнее положение связано с тем, что каждая зона мозга, участвующая в обеспечении функциональной системы, ответственна за свой фактор, и его устранение приводит к тому, что нормальное осуществление функции становится невозможным. Приведенный выше пример построения произвольного движения и видов его нарушения при локальных поражениях мозга отчетливо это иллюстрирует.
Описанные закономерности построения и распада функциональных систем позволяют нейропсихологу выяснить, какие именно факторы включает в свой состав сложная психическая деятельность и какие именно участки мозга составляют ее нервную основу.
Оба эти вопроса могут быть решены только при сопоставлении всех симптомов, которые отмечаются при наличии одного, строго локализованного очага в коре головного мозга (или в подкорковых образованиях), с одной стороны, и тщательного анализа характера нарушений данной системы при различных по локализации мозговых поражениях — с другой.
Остановимся на иллюстрации этого положения.
84
Как уже было сказано ранее, для успешного осуществления сложного движения нужна его четкая пространственная организация, иначе говоря, четкое построение движения в системе пространственных координат. Это условие обеспечивается третичными (зрительно-кинестетически-вестибулярными) отделами теменно-затылочной коры, несоблюдение этого условия приводит к распаду пространственно-организованного движения. Возникает, однако, естественный вопрос: какие другие виды психической деятельности нарушаются при поражении тех же теменно-затылочных отделов мозга, обеспечивающих функцию пространственной организации процессов? Если мы получим ответ на этот вопрос и если мы сможем выделить одну группу процессов, страдающих при такой локализации очага, и другую группу процессов, которая при данном патологическом очаге остается сохранной, мы значительно приблизимся к выяснению того, в какие именно виды психической деятельности входит пространственный фактор, имеющий прямую связь с указанными теменно-затылочными отделами мозговой коры.
Факты, показывающие, что любой ограниченный корковый очаг поражения действительно нарушает протекание одних психических процессов, оставляя другие процессы в сохранности (явление, названное американским нейропсихологом Г.Л.Тэйбером принципом двойной диссоциации функций), в изобилии содержатся в материалах нейропсихологических исследований.
Так, ограниченный очаг в теменно-затылочной (или нижнетеменной) области левого полушария, приводящий к нарушению пространственной организации восприятия и движения, неизбежно вызывает и другие симптомы: такие больные, как правило, не могут ориентироваться в положении стрелок на часах или в координатах на географической карте; они не могут ориентироваться в плане клинического отделения, где они находятся; не могут решить даже относительно несложные арифметические примеры, смешивая, например, при вычитании из двузначного числа с переходом через десяток направление операций (решая пример 31 — 7, они правильно выполняют первый этап — от 30 отнимают 7 и получают 23, но затем не знают, направо или налево надо отложить оставшуюся единицу и какой окончательный ответ — «22» или «24» является правильным); наконец, они начинают испытывать серьезные затруднения в понимании ряда грамматических структур, включающих в свой состав известные логические отношения (например, «брат отца» и «отец брата», «весна после лета» или «лето после весны»), в то время как понимание других, более простых грамматических структур остается сохранным.
Однако указанный очаг не приводит к нарушениям таких процессов, как плавная речь, узнавание или воспроизведение музыкальных мелодий, смена последовательных элементов движения и т.д.
85
Все это показывает, что первая группа отмеченных выше процессов включает в свой состав «пространственный» фактор, в то время как вторая группа процессов этого фактора не включает и поэтому остается сохранной при поражении теменно-затылочных отделов коры.
Совершенно обратная картина наблюдается при локальных поражениях височной (слуховой) области коры. Поражения с такой локализацией, как мы увидим далее, приводят к нарушению организации слухового восприятия и делают невозможной организацию звуковых сигналов в последовательную («сукцессивную») структуру. Именно в силу этого больные с такими поражениями оказываются не в состоянии четко воспринимать обращенную к ним речь и удерживать ее следы; плавная избирательная речь, как и слухоречевая память, оказывается у них серьезно нарушенной. Однако такие функции, как ориентировка в пространстве, пространственная организация движений, счетные операции, понимание определенных логико-грамматических отношений, остаются у них, как правило, сохранными.
Все это с полной отчетливостью показывает, что тщательный нейропсихологический анализ синдрома и той двойной диссоциации, которая возникает при локальных поражениях мозга, позволяет приблизиться к структурному анализу самих психологических процессов и выделить те факторы, которые входят в одни группы психических процессов и не входят в другие.
Как мы увидим далее, это позволяет вплотную подойти к решению вопроса о внутреннем составе психологических процессов, который оставался неразрешимым в условиях обычного психологического исследования, и, таким образом, разделить, казалось бы, сходные психологические процессы и сблизить, казалось бы, различные формы психической деятельности.
Проиллюстрируем это положение двумя примерами.
Для непредвзятого наблюдателя музыкальный и речевой слух могут казаться двумя вариантами одного и того же психологического процесса. Однако наблюдения над больными с ограниченными очаговыми поражениями мозга показывают, что разрушение определенных участков левой височной области приводит к выраженному нарушению речевого слуха (делая различение близких звуков речи совершенно недоступным), но сохраняет музыкальный слух неповрежденным. В наших публикациях есть описание одного выдающегося композитора, который после кровоизлияния в левую височную область перестал различать звуки речи и понимать обращенную к нему речь, но продолжал создавать блестящие музыкальные произведения (А. Р. Лурия, Л. С. Цветкова, Д. С. Футер, 1965).
Это означает, что столь близкие, казалось бы, психические процессы, как музыкальный и речевой слух, не только включают в свой состав разные факторы, но и опираются на работу различных мозговых зон.
86
Нейропсихологии известны также примеры, показывающие внутреннюю близость, казалось бы, полностью различных психологических процессов.
Вряд ли кто-нибудь сразу согласится с тем, что столь различные психологические процессы, как ориентировка в пространстве, счет и понимание сложных логико-грамматических структур, имеют принципиально общие звенья, которые позволяют объединить их в одну группу.
Однако поражение теменно-затылочных (нижнетеменных) отделов левого полушария почти неизбежно приводит к нарушению всех этих процессов, и больной с подобной локализацией очага не только испытывает заметные трудности в пространственной ориентировке, но и обнаруживает грубейшие дефекты в счете и в понимании сложных логико-грамматических структур.
Это показывает, что все указанные, казалось бы, столь различные функции включают общий фактор, и выделение этих общих факторов способствует гораздо более глубокому анализу структуры психологических процессов.
Нетрудно увидеть, что синдромный анализ позволяет не только уточнить вопрос о мозговой организации сложных психических процессов, но и вплотную подойти к их внутреннему строению.
Итак, каждая сознательная психическая деятельность представляет собой сложную функциональную систему, которая может нарушаться в разных звеньях и страдать при различных по локализации поражениях головного мозга (но по-разному); в ее состав включены различные факторы, изучение которых открывает новые пути для нейрофизиологического анализа внутреннего строения психических процессов.
Наблюдения над изменениями психических процессов, возникающими при локальных поражениях мозга, могут действительно стать одним из наиболее существенных источников наших знаний о мозговой организации психической деятельности. Однако правильное использование этого метода возможно лишь при условии отказа от концепции прямой локализации психических процессов в коре головного мозга и замены этой классической задачи другой, требующей анализа того, как именно меняется психическая деятельность при различных локальных поражениях мозга и какие именно факторы вносит каждый из аппаратов головного мозга в построение сложных форм психической деятельности.
Эта новая задача определяет генеральный путь нейропсихологии как учения о мозговой организации психических процессов человека.
Глава III
ТРИ ОСНОВНЫХ ФУНКЦИОНАЛЬНЫХ БЛОКА МОЗГА
Мы уже говорили, что психические процессы человека являются сложными функциональными системами и что они не локализованы в узких, ограниченных участках мозга, а осуществляются при участии сложных комплексов совместно работающих мозговых аппаратов, каждый из которых вносит свой вклад в организацию этой функциональной системы. Вот почему становится необходимым выяснить, из каких основных функциональных единиц состоит мозг человека, как построена и какую роль играет каждая из них в осуществлении сложных форм психической деятельности.
Можно выделить три основных функциональных блока, или три основных аппарата мозга, участие которых необходимо для осуществления любого вида психической деятельности. С некоторым приближением к истине их можно обозначить как: 1) блок, обеспечивающий регуляцию тонуса и бодрствования; 2) блок получения, переработки и хранения информации, поступающей из внешнего мира; 3) блок программирования, регуляции и контроля психической деятельности.
Каждый из этих основных блоков имеет иерархическое строение и состоит по крайней мере из надстроенных друг над другом корковых зон трех типов: первичных (или проекционных), куда поступают импульсы с периферии или откуда направляются импульсы на периферию, вторичных (или проекционно-ассоциативных), где происходит переработка получаемой информации или подготовка соответствующих программ, и, наконец, третичных (или зон перекрытия), которые являются наиболее поздно развивающимися аппаратами больших полушарий и которые у человека обеспечивают наиболее сложные формы психической деятельности, требующие совместного участия многих зон мозговой коры.
Рассмотрим строение и функциональные особенности каждого из этих блоков головного мозга в отдельности.