
Регуляция нервной системой








Секреция катехоламина адреналина из мозгового вещества надпочечников контролируется симпатической нервной системой. При этом никакие сдвиги метаболизма, вызванные адреналином, не влияют на его секрецию.
71. Основные механизмы регуляции метаболизма: механизмы действия гормонов, изменяющих активность ферментов, количество ферментов в клетке, проницаемость клеточных мембран. Омрммдгиафкфвкпкм
Стр 378
Регуляция обмена белков, жиров и углеводов контролируется действием гормонов, включая инсулин, глюкагон, адреналин и кортизол. Изменения уровней этих гормонов позволяет организму запасать энергию, когда пища присутствует в достаточных количествах, или делать доступными для использования энергетические запасы во время “кризов выживания”, таких как голодание, тяжелые повреждения, или реакция “борьбы или бегства”. Учитывая распределение энергетических ресурсов (табл.1), не удивительно, что именно печень, скелетные мышцы и жировая ткань в первую очередь являются объектами воздействия вышеперечисленных гормонов. Конечными (интегральными) показателями влияния указанных гормонов на обмен углеводов, жиров и белков является их влияние на уровни глюкозы, свободных жирных кислот (СЖК) и аминокислот (АК) в плазме крови

Влияние на обмен углеводов
Инсулин:
1) ингибирует глюконеогенез в печени, повышая уровень внутриклеточного регулятора фруктозо-2,6-бисфосфата. [Замечание: наи-
более важные эффекты данного внутриклеточного регулятора обмена
углеводов – активация фосфофруктокиназы-1 и ингибирование фрук-
тозо-1,6-бисфосфатазы]. Кроме того, инсулин ингибирует глюкозо-6-
фосфатазу и синтез фосфоенолпируваткарбоксикиназы (ключевые
ферменты глюконеогенеза);
2) уменьшает распад гликогена (инактивируя киназу фосфорила-
зы) и повышает его синтез (активируя гликогенсинтазу);
3) интенсифицирует реакции гликолиза (без образования лакта-
та!), повышая активность и количество ключевых ферментов – глюко-
и гексокиназы, фосфофруктокиназы и пируваткиназы;
4) повышает активность ферментов пентозофосфатного пути –
одного из основных генераторов молекул НАДФН, необходимых для
синтеза жирных кислот;
5) в мышечной и жировой ткани инсулин усиливает поступление
глюкозы в клетки (через увеличение числа GLUT-4).
Глюкагон
Активация аденилатциклазы и повышение концентрации цАМФ обуславливает усиление распада гликогена в печени. В отличие от адреналина глюкагон не влияет на гликогенолиз в скелетных мышцах. Высокий уровень цАМФ обуславливает
активацию транскрипции гена фосфоенолпируваткарбоксикиназы –
ключевого фермента глюконеогенеза. Кроме того, накопление цАМФ
приводит к активации фруктозо-1,6-бисфосфатазы – второго ключево-
го фермента глюконеогенеза.
В печени гормон угнетает синтез белков и облегчает их катабо-
лизм. Высвобождающиеся аминокислоты используются в реакциях
глюконеогенеза.
Таким образом, центральный эффект глюкагона – гипергликемия
– обеспечивается двумя механизмами:
1. Быстрый – распад гликогена,
2. Медленный – стимуляция глюконеогенеза.
Адреналин.
Главным местом метаболического действия катехоламинов яв-
ляются печень, мышечная и жировая ткани. Главными метаболически-
ми эффектами, опосредуемыми различными адренорецепторами явля-
ются:
1. Стимуляция распада гликогена до глюкозы (гликогенолиза) в
печени (посредством активации a1 и b2-адренорецепторов) и мышцах
(через b2-рецепторы). В быстросокращающихся скелетных мышцахглюкоза метаболизируется до лактата. В тоже время глюкоза, образуемая в печени, поступает в кровоток. Это приводит к повышению
уровня глюкозы и лактата в плазме крови.
2. Активация глюконеогенеза (через b2-рецепторы) из лактата и
глицерола.
7. Ингибирование секреции инсулина (через a2-рецепторы). Это
снижает утилизацию глюкозы скелетными мышцами и жировой тканью, сберегая ее для ЦНС.
72. Инсулин: строение, синтез, механизм действия, метаболические эффекты. Иссмдмэ
Стр 400
73. Характеристика нарушений обмена веществ при сахарном диабете. Биохимические механизмы развития осложнений сахарного диабета. Хновпсдбмросх стр 407-409
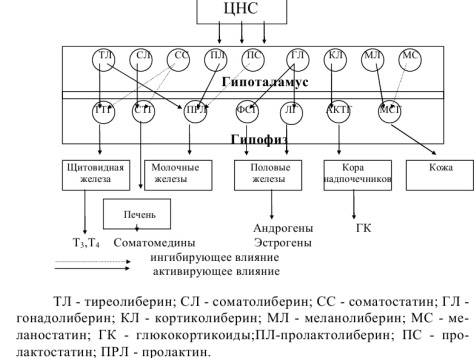
74. Центральная регуляция эндокринной системы: гормоны гипоталамуса и тропные гормоны гипофиза. Црэсггитгг
Регуляция синтеза и секреции тропных гормонов гипофиза осуществляется путем влияния гипоталамических нейрогормонов, поступающих в гипофиз через портальную систему сосудов.
Классификация гипоталамических нейрогормонов основана на их способности стимулировать (либерины) или ингибировать (статины) высвобождение соответствующего гормона гипофиза.
К первой группе относятся кортиколиберин – гормон, высвобождающий АКТГ; тиреолиберин – гормон, высвобождающий тиреотропный гормон; соматолиберин – гормон, высвобождающий соматотропный гормон;
гонадолиберин – гормон, высвобождающий фолликулостимулирующий и лютеинизирующий гормоны;
пролактолиберин – гормон, высвобождающий пролактин; мела-
нолиберин – гормон, высвобождающий меланоцитстимулирующий гормон.
Ко второй группе относятся соматостатин – гормон, ингибирующий выделение соматотропина;
пролактостатин – гормон, ингибирующий выделение пролактина;
меланостатин – гормон, ингибирующий выделение меланоцитстимулирующего гормона.
К гипоталамическим гормонам можно отнести также антидиуретический гормон (вазопрессин) и окситоцин, синтезирующиеся в ядрах гипоталамуса и транспортируемые затем в заднюю долю гипофиза. Все гипоталамические гормоны, за исключением пролактостатина, – это вещества пептидной природы. Установлено точное строение пяти гипоталамических нейрогормонов: тиреолиберина, гонадолиберина, соматостатина, кортиколиберина и соматолиберина. Эти гормоны состоят соответственно
из 3, 10, 14, 41 и 44 аминокислотных остатков. Точное строение остальных гипоталамических гормонов полностью не установлено. Гипоталамический фактор, ингибирующий синтез пролактина (пролактостатин), идентифицирован как дофамин.
Гипофизарные гормоны представляют собой группу белковопептидных гормонов. В передней доле гипофиза вырабатываются адренокортикотропный гормон (АКТГ), тиреотропный гормон (ТТГ), лютеинизирующий гормон (ЛГ), фолликулостимулирующий гормон (ФСГ), пролактин (ПРЛ) и соматотропный гормон (СТГ) или гормонроста. В промежуточной доле гипофиза синтезируется меланоцитстимулирующий гормон (МСГ), а в задней доле накапливаются антидиуретический гормон (АДГ) и окситоцин.
Гипоталамо-гипофизарные гормоны очень быстро исчезают из крови, так как не имеют специфических белков-переносчиков. Период их полураспада составляет несколько минут.
Инактивация (распад) этих гормонов происходит в печени под действием специфических пептидаз.
Соматотропный гормон (гормон роста) наиболее важный гормон для достижения нормального роста. СТГ вырабатывается в передней доле гипофиза и представляет собой белок, состоящий из 191 аминокислотного остатка.
Секреция гормона роста возрастает во сне, при физических упражнениях, стрессе и гипогликемии, а также стимулируется соматолиберином гипоталамуса. Соматостатин и соматомедины ингибируют секрецию гормона. Гормон роста также ингибирует свою секрецию через стимуляцию высвобождения соматостатина из гипоталамуса.
75. Гормон роста: строение, регуляция выработки, биологическая роль. Соматомедины. Проявления избытка и недостатка гормона. Грсрвбрспиинг
Строение
Представляет собой полипептид, включающий 191 аминокислоту с молекулярной массой 22 кДа и периодом полураспада 20-25 мин.
Синтез
Осуществляется в ацидофильных клетках гипофиза – подкласс соматотрофов с волнообразной секрецией и пиком каждые 20-30 мин.