
III. Биологическое окисление.








Биологическое окисление – это совокупность реакций окисления, протекающих в живом организме.
Лавуазье сравнивал биологическое окисление с «медленным горением», но это ограниченная аналогия, так как биологическое окисление:
а) протекает при низкой температуре,
б) протекает без появления пламени,
в) протекает в присутствии воды.
Существует несколько теорий биологического окисления:
1. Теория “активации” кислорода (Бах)
Образуются пероксиды: а) О=О®–О–О–
б) –О–О– + S ® SOO [треугольник, в углах которого три указанные буквы; S = субстрат; над стрелочкой реакции надпись “оксигеназы”]
в) SOO + S’ ® SO + S’O [фермент пероксидаза]
Эта теория не объясняет окисление в животных тканях.
2. Теория активирования водорода (Палладин)
В клетках животных окисление идет благодаря дегидрированию:
А·Н2 + Ко® А + Ко·Н2 [фермент дегидрогеназа]
Ко·Н2 + ½О2 ® Ко + Н2О
3. Современные представления (Палладин и Бах)
Биологическое окисление – это процесс переноса электронов. Если акцептором электронов выступает молекулярный кислород, то его называют “тканевым дыханием”:
RH2 ® R + 2H+ + 2e--
2H+ + 2e-- + ½ O2 ® H2O + 210 кДж
Биологическое окисление – многоступенчатый полиферментативный процесс, заключающийся в многократной передаче протонов и электронов по цепи ферментов. При этом химическая энергия выделяется небольшими порциями (постепенно, без взрывов).
Дыхательная цепь (ДЦ)
(или Цепь Переноса Электронов – ЦПЭ, или Электрон-Транспортная Цепь – ЭТЦ)
ДЦ – это конвейер по переносу электронов и протонов от восстановленного субстрата к кислороду.
Компоненты ДЦ:
1. Пиридинзависимые ДГ (НАД-, НАДФ-зависимые)
Рабочая часть – витамин РР (никотинамид)
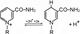
НАД + 2Н+ + 2е ↔ НАД·Н2
2. ФАД-зависимые ДГ (кофермент в ДЦ – ФМН, а акцептор электронов непосредственно от субстрата – ФАД. Рабочая часть – изоалоксазин.
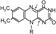
[При восстановлении к атомам азота при двойных связях, отмеченных стрелками, присоединяется по атому водорода, а двойная связь перемещается на общую грань колец В и С.]
3. Убихинон (Ko Q). Обладает о/в-свойствами благодаря кето-енольной таутомерии.
4. Цитохромы. Относятся к гемопротеинам, содержат атомы железа, переход степени (2↔3) окисления которого и обеспечивает транспорт электронов (протоны ими не транспортируются !!! ).
В ДЦ цитохромы расположены в следующей последовательности: b - c1 - c - a - a3 .
Совокупность цитохромов b и c1 называют КоQH-дегидрогеназой, т.к. они отщепляют атом водорода от убихинона (KoQ).
Цитохромы а и а3 – цитохромоксидазой (т.к. способствуют переносу электронов на молекулярный кислород).
Функционирование ДЦ
Субстрат·Н2 → НАД → ФМН → КоQ → 2b → 2c1 → 2c → 2a → 2a3 → O2 .
[До KoQ включительно переносятся 2 протона и 2 электрона, а по цепи цитохромов – только 2 электрона]
Существует и укороченная ДЦ, в которой субстрат окисляется ФАД-зависимой ДГ, отдающей затем 2 протона и 2 электрона непосредственно на убихинон.
Необходимо отметить, что АТФ выделяется на этапах: НАД→ФМН (в укороченной ДЦ эта молекула АТФ не выделяется!), b→c1 , a→a3 .
Вообще, молекула АТФ синтезируется если разница потенциалов между соседними компонентами цепи превышает 0,2 В, т.е. может выделиться энергия не менее 50 кДж/моль.